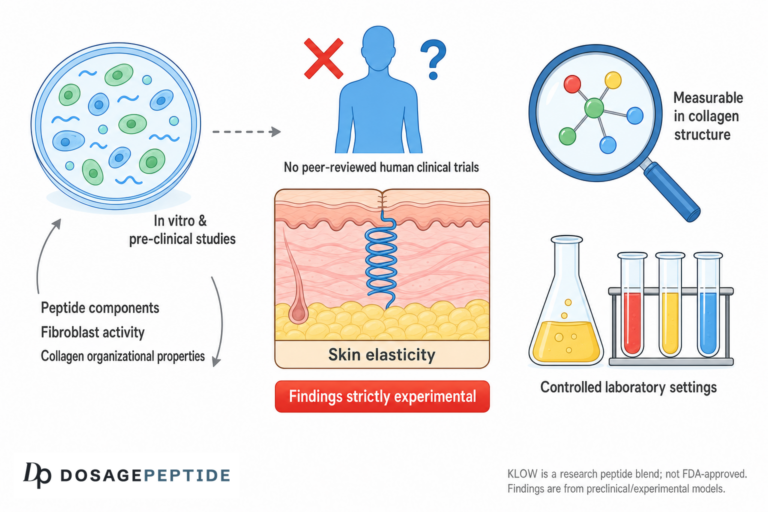
The question in the title carries a quiet assumption that deserves to be examined before anything else: that “studies” have looked at KLOW peptides and energy metabolism at all, and that the two belong together in a sentence. They largely do not. KLOW is a fixed, multi-peptide research blend — a lyophilized combination of GHK-Cu, BPC-157, TB-500, and KPV, sold to laboratories as a tissue-repair reagent rather than a metabolic agent.1 None of its four components was designed to modulate how cells generate, store, or spend energy, and there are no clinical trials — none — testing the KLOW blend as a whole against any endpoint, let alone an energy-metabolism endpoint such as resting metabolic rate, mitochondrial respiration, glucose disposal, or exercise capacity in humans. The honest starting point is that KLOW and “energy metabolism” are joined mostly by marketing and inference, not by data.
That does not make the question worthless. It makes it an open research question, and a genuinely interesting one, because each of KLOW’s ingredients touches biology that is at least adjacent to energy metabolism: copper is an obligatory cofactor for the mitochondrial enzyme that finishes cellular respiration; angiogenesis determines how much oxygen and fuel a tissue can receive; chronic inflammation imposes a measurable metabolic cost that anti-inflammatory signaling might relieve. So there are plausible, mechanistic threads one can pull. The task of this article is to pull them honestly — to distinguish a real but indirect biochemical link from a demonstrated metabolic effect, and to be explicit that the distance between “copper participates in ATP synthesis” and “this peptide blend improves your energy metabolism” is enormous and, at present, unbridged by evidence.
This piece is written for researchers and scientifically literate readers who want a truthful map rather than a sales pitch. We will define what KLOW actually is and where it came from, unpack what “energy metabolism” means physiologically, examine each component’s biology and its real (usually indirect) relationship to metabolism, survey the actual evidence and the size of the human-trial gap, compare KLOW with compounds that have been studied for metabolic endpoints, cover research handling, safety, and regulatory status, and answer the questions people actually ask. Throughout, one rule governs: KLOW is an unapproved research blend, its individual peptides are themselves largely investigational or preclinical, and nothing here should be read as evidence that KLOW treats fatigue, boosts metabolism, or improves any disease of energy balance.
What KLOW Actually Is: A Four-Peptide Repair Blend
KLOW is not a single molecule but an acronym-style trade name for a specific combination of four research peptides, most commonly supplied as an 80 mg lyophilized vial with a fixed ratio: 50 mg GHK-Cu, 10 mg BPC-157, 10 mg TB-500, and 10 mg KPV.1 The name is a loose contraction of its ingredients (K for KPV, and the “LOW” evoking the “GLOW” family of skin-and-repair blends to which it is closely related). It is best understood as a “GLOW-plus-KPV” formulation: the popular GLOW blend pairs GHK-Cu, BPC-157, and TB-500 for skin and soft-tissue research, and KLOW adds the anti-inflammatory tripeptide KPV on top. For readers exploring the sibling formulation, the site’s coverage of KLOW peptides in angiogenesis and tissue repair traces the blend’s primary, intended research context, which is regeneration — not metabolism.
Each ingredient carries its own history and its own body of (mostly preclinical) literature. GHK-Cu is the copper-bound tripeptide glycyl-L-histidyl-L-lysine, discovered by Loren Pickart in human plasma in the 1970s and studied for decades as a modulator of tissue remodeling and gene expression.1 Its most-cited modern review, drawing on Broad Institute Connectivity Map data, reports that GHK can shift the expression of a large fraction of human genes toward patterns associated with tissue repair, up-regulating antioxidant and wound-healing programs while helping to reset age-related transcriptional changes.1 A companion review details how GHK-Cu stimulates fibroblast function, collagen and glycosaminoglycan synthesis, and balanced matrix remodeling through coordinated up-regulation of matrix metalloproteinases and their inhibitors.2 The copper atom it carries is not incidental — it is central to why GHK-Cu has any conceivable connection to energy metabolism at all, a point we develop below.
BPC-157 is a synthetic pentadecapeptide (fifteen amino acids) whose sequence is derived from a partial fragment of a protective protein identified in human gastric juice. In rodent models it has been reported to accelerate the healing of tendon, muscle, gut, and other tissues, and much of its proposed mechanism centers on angiogenesis and the nitric-oxide system.3 Cell and vessel studies indicate that BPC-157 up-regulates VEGF receptor-2 (VEGFR2) and drives downstream Akt–eNOS signaling to promote endothelial migration and new-vessel formation,3 and separate work implicates a Src–caveolin-1–eNOS pathway in its effects on vascular tone.4 Comprehensive reviews frame BPC-157’s pleiotropic actions as fundamentally tied to modulating angiogenesis and nitric-oxide biology.5 Readers wanting the fuller primer can consult the site’s explainer on what BPC-157 is and how it is studied. Crucially, none of this is metabolic-rate biology; it is repair and perfusion biology.
TB-500 is a synthetic peptide corresponding to the active region of thymosin β4, a naturally occurring 43-amino-acid protein whose principal molecular job is to bind monomeric G-actin and regulate the actin cytoskeleton.6 Through that actin-regulating activity and related signaling, thymosin β4 promotes cell migration, angiogenesis, and wound repair, and it has a notable preclinical cardiac literature: a landmark study showed that thymosin β4 activates integrin-linked kinase and Akt to enhance cardiomyocyte survival and cardiac repair after injury.7 As with BPC-157, the compound’s documented biology is about building and repairing tissue, not about setting metabolic rate. The site’s discussion of TB-500 in chronic inflammatory conditions surveys where the preclinical interest actually lies.
KPV is the smallest of the four: a tripeptide of lysine, proline, and valine corresponding to the C-terminal residues (11–13) of α-melanocyte-stimulating hormone (α-MSH). It retains much of α-MSH’s anti-inflammatory activity while lacking the parent hormone’s pigmentary and, importantly, its melanocortin-receptor-mediated effects. In murine models of inflammatory bowel disease, KPV reduced intestinal inflammation,8 and targeted oral-delivery studies have shown it can attenuate experimental colitis at very low doses.9 KPV’s parentage in α-MSH is the second thread — alongside GHK-Cu’s copper — that gives KLOW even a tenuous claim on the word “metabolism,” because the melanocortin system from which KPV derives is deeply involved in energy homeostasis. Whether KPV itself carries any of that metabolic activity is a separate and largely unstudied question we return to below.
The single most useful habit when thinking about KLOW is to resist treating the blend as though it were a designed drug with a unified mechanism. It is four different molecules, from four different research lineages, combined at a fixed ratio for a repair-oriented rationale. Any claim about KLOW “and energy metabolism” is really a claim about some emergent or additive metabolic effect of four peptides that were never tested together for that purpose. That is a strong claim, and the burden of proof for it has not been met.
Reframing the Question: What “Energy Metabolism” Actually Means

To judge whether KLOW could plausibly affect energy metabolism, we have to be precise about what that phrase denotes, because in popular usage it collapses several distinct biological systems into a vague notion of “feeling energetic.” Scientifically, energy metabolism refers to the network of processes by which cells extract chemical energy from nutrients and convert it into usable forms — principally adenosine triphosphate (ATP) — and by which the whole organism balances energy intake against expenditure.
At the cellular level, the core machinery is mitochondrial. Glucose and fatty acids are broken down through glycolysis and β-oxidation into acetyl-CoA, which feeds the tricarboxylic-acid cycle; the electrons harvested there flow down the mitochondrial electron transport chain, and the resulting proton gradient drives ATP synthase. The terminal, rate-limiting enzyme of that chain is cytochrome c oxidase (Complex IV), which reduces molecular oxygen to water — and it cannot function without copper. Copper is an obligatory catalytic cofactor of cytochrome c oxidase, forming the CuA and CuB centers that shuttle electrons to oxygen, and its delivery to the enzyme requires a dedicated set of mitochondrial chaperone proteins.10 Defects in that copper-delivery pathway cause fatal deficiencies of cytochrome c oxidase activity, underscoring how tightly copper and cellular ATP production are linked.10 This is the most direct biochemical bridge between anything in KLOW and energy metabolism, and it runs entirely through the copper carried by GHK-Cu.
At the whole-body level, energy metabolism is governed by an endocrine and neural control network: insulin and glucagon manage substrate storage and release; thyroid hormone sets basal metabolic rate; catecholamines and the sympathetic nervous system drive acute energy mobilization and thermogenesis; and the hypothalamic melanocortin system integrates signals of energy stores (leptin, insulin) to regulate food intake and energy expenditure. The melanocortin-4 receptor in particular is a critical coordinator of mammalian energy homeostasis and body weight.12 Because KPV is an α-MSH fragment, the melanocortin system is the second bridge worth examining — though, as we will see, KPV appears to have been specifically shorn of the receptor-binding activity through which α-MSH exerts its metabolic effects.
The practical upshot of this framing is a checklist. For KLOW to “affect energy metabolism” in any meaningful, measurable sense, at least one of its components would need to demonstrably change mitochondrial respiration, substrate handling (glucose or lipid disposal), basal or activity-related energy expenditure, or the endocrine signals that set those parameters — and that change would need to be shown, ideally, in a living organism and eventually in humans. What we have instead is a set of indirect, mechanistically plausible possibilities, none of which has been tested for the blend and few of which have been tested even for the individual peptides against a genuine metabolic endpoint. Keeping the checklist in mind is what separates responsible analysis from wishful extrapolation.
The Copper Connection: GHK-Cu, Mitochondria, and an Indirect Metabolic Link
If any part of KLOW has a legitimate claim on energy metabolism, it is GHK-Cu — and the claim rests almost entirely on the copper atom it carries rather than on any demonstrated metabolic action of the peptide itself. The logic is straightforward and biochemically sound: cytochrome c oxidase, the enzyme that completes cellular respiration and enables the bulk of ATP production, is copper-dependent, and adequate copper delivery is essential for mitochondrial energy output.10 GHK-Cu is, among other things, a copper-delivery vehicle — the peptide binds copper(II) with high affinity and can exchange it with cellular copper pools. In principle, a copper-carrying tripeptide that improves copper availability could support the biogenesis and function of copper-dependent enzymes, including cytochrome c oxidase.
That is a real mechanistic thread, and it should not be dismissed. But it must be handled with care, because plausibility is not proof, and the gap between the two is where most overstatement lives. Several honest caveats apply. First, the primary GHK-Cu literature documents its effects on gene expression, tissue remodeling, wound healing, antioxidant defense, and skin regeneration — not on measured mitochondrial respiration or systemic energy expenditure.12 The gene-modulation review notes GHK’s influence on a very broad set of cellular pathways, and some of those touch mitochondrial and antioxidant biology, but “influences genes related to mitochondrial function” is a long way from “increases ATP production” in a way a person would experience as energy.1 Second, copper is a double-edged nutrient: too little impairs respiration, but excess free copper is pro-oxidant and cytotoxic, and the body regulates copper tightly. Supplying copper via a peptide is not automatically beneficial, and in a copper-replete system it confers no obvious metabolic advantage.
Third, and most important for the KLOW question specifically, the dose and route matter enormously. A research vial delivering copper as part of a GHK-Cu blend is a fundamentally different proposition from a controlled study of mitochondrial copper loading, and no one has demonstrated that GHK-Cu, at any dose used in these blends, measurably increases cytochrome c oxidase activity or ATP output in a living human. The mechanistic story is best summarized as: copper is genuinely essential for cellular energy production; GHK-Cu genuinely carries copper; therefore GHK-Cu could, in a copper-limited context, support the machinery of energy metabolism — but this remains a hypothesis about a permissive, cofactor-level role, not evidence of an energizing effect. It is the difference between saying “iron is needed to make hemoglobin” and “taking iron will give you more energy,” which is only true in the specific case of deficiency.
GHK-Cu’s better-documented biology — the reason it anchors repair blends — lies in regeneration and gene modulation, as detailed in research on GHK-Cu in chronic non-healing wounds. That regenerative activity has an oblique metabolic dimension worth naming: healthy, well-vascularized, low-inflammation tissue is metabolically more efficient than damaged, hypoxic, inflamed tissue. To the extent GHK-Cu supports tissue quality, it may indirectly support the conditions under which cells metabolize efficiently. But this is a downstream, contextual benefit of repair, not a primary metabolic action, and framing it as “GHK-Cu boosts energy metabolism” would badly overstate what the data show.
BPC-157, Angiogenesis, and the Delivery of Fuel and Oxygen
BPC-157’s relationship to energy metabolism is even more indirect than GHK-Cu’s, and it runs through the circulatory system rather than the mitochondrion. The peptide’s best-characterized action is pro-angiogenic: in endothelial-cell and vessel studies it up-regulates VEGFR2 and activates the VEGFR2–Akt–eNOS axis, increasing nitric-oxide production and promoting the migration and organization of endothelial cells into new vessels.3 Additional work implicates a Src–caveolin-1–eNOS pathway in its modulation of vascular tone,4 and reviews consistently place angiogenesis and nitric-oxide signaling at the center of its pleiotropic, tissue-protective effects.5
Why might that matter for metabolism? Because energy metabolism is ultimately constrained by delivery. Mitochondria can only oxidize the oxygen and substrate that reach them, and that delivery depends on capillary density and blood flow. In tissues where perfusion is the limiting factor — a healing wound, an ischemic muscle, a poorly vascularized graft — improving angiogenesis can improve the local supply of oxygen and nutrients and, with it, the tissue’s capacity for aerobic energy production. This is a genuine and important principle. But it is a principle about local perfusion in compromised tissue, not about whole-body energy metabolism in a healthy organism, and the distinction is decisive.
In a person with normal vasculature, capillary supply is not the bottleneck on energy metabolism, and there is no reason to expect a pro-angiogenic peptide to raise metabolic rate, improve glucose disposal, or increase ATP availability systemically. Moreover, all of BPC-157’s angiogenic data come from cell culture and rodent models; the peptide is not approved for human use for any indication, and the pro-angiogenic property that could be helpful in a healing wound is the same property that raises legitimate caution in other contexts, since indiscriminate angiogenesis is not universally desirable. The honest reading is that BPC-157 could, in specific pathological settings, improve the perfusion side of the energy-supply equation, but that this is a repair-and-delivery mechanism rather than a metabolic one, and it provides no basis for claiming that KLOW “improves energy metabolism” in the everyday sense the phrase implies.
There is also a physiological subtlety that cuts against naive optimism. Angiogenesis and tissue repair are themselves energetically expensive: building new vessels and remodeling matrix consumes ATP and substrate. In the short term, an actively repairing tissue may have a higher, not lower, local energy demand. So even the local metabolic effect of BPC-157-driven angiogenesis is not simply “more energy available”; it is a reallocation of energy toward construction. This is a useful reminder that “affecting energy metabolism” is not synonymous with “giving you energy” — a conflation that runs through nearly all consumer-facing peptide marketing.
TB-500 / Thymosin β4: Actin, Repair, and Metabolic Cost
TB-500’s connection to energy metabolism is the most tenuous of the four, and being clear about that is part of writing honestly. The peptide’s defining molecular activity is the sequestration of monomeric actin and the regulation of the actin cytoskeleton, an activity inherited from its parent protein thymosin β4.6 Through cytoskeletal regulation and associated signaling, thymosin β4 promotes cell migration, angiogenesis, survival, and wound healing across many tissue types,6 and in cardiac injury models it activates integrin-linked kinase and Akt to protect cardiomyocytes and support repair.7
None of this is energy-metabolism biology in any direct sense. Actin dynamics, cell migration, and cardioprotection are cellular-architecture and survival processes. One can construct indirect links — the Akt signaling that thymosin β4 engages is part of a broad network that also intersects with metabolic regulation, and preserving cardiomyocytes after ischemia could, trivially, preserve the heart’s capacity to do metabolic work — but these are inferences several steps removed from any measured metabolic outcome. There is no body of work showing that TB-500 changes mitochondrial respiration, glucose uptake, fat oxidation, or energy expenditure. Its cardiac literature, moreover, illustrates the broader translational challenge: despite encouraging preclinical results, human trials of thymosin β4-related approaches in cardiac repair have produced mixed outcomes, a reminder that animal findings frequently fail to reproduce in people.
The one metabolically flavored point worth making about TB-500 is the same one that applies to BPC-157: regeneration has an energy cost. A migrating, proliferating, repairing cell is doing energetically demanding work, and TB-500’s pro-migratory, pro-repair activity operates within that expensive program. If anything, the compound’s biology is about directing and enabling the use of energy for construction, not about increasing the supply of energy or improving the efficiency of its production. For the KLOW “energy metabolism” question, TB-500 is best described as a repair-and-motility peptide whose relevance to metabolism is speculative and, at present, entirely unsupported by direct data. Its genuine research interest lies elsewhere, in tissue and possibly cardiac regeneration, where the evidence — while still preclinical and mixed — is at least on-topic.
KPV, Inflammation, and the Metabolic Tax of Chronic Immune Activation
KPV brings the most conceptually interesting — and most double-edged — connection to energy metabolism, for two separate reasons: its documented anti-inflammatory activity, and its parentage in the metabolically active hormone α-MSH.
Start with the anti-inflammatory route, which is the better supported. KPV is a potent inhibitor of NF-κB signaling, the master transcriptional regulator of inflammatory gene expression, and it reduces the production of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6. In murine inflammatory-bowel-disease models it attenuates intestinal inflammation,8 and low-dose targeted-delivery studies confirm efficacy in experimental colitis.9 The metabolic relevance here is real but indirect: chronic inflammation imposes a genuine metabolic burden. Systemic inflammatory activation raises resting energy expenditure in a wasteful way, drives insulin resistance, impairs mitochondrial function, and promotes catabolism — the constellation seen in conditions from chronic infection to obesity-associated metabolic disease. In principle, a compound that dampens chronic, low-grade inflammation could relieve some of this “inflammatory tax” and thereby improve metabolic efficiency and insulin sensitivity.
That is a legitimate hypothesis, and it is the strongest single argument for KLOW having any metabolic relevance. But it must be stated with matching caveats. The link is indirect (inflammation reduction → metabolic improvement), it has been demonstrated for KPV only in inflammation models rather than metabolic ones, and the metabolic benefit of anti-inflammatory action is context-dependent — it is most plausible in states of pathological inflammation and largely irrelevant in a healthy, non-inflamed organism. KPV has not been tested as a metabolic agent, has no human metabolic-outcome data, and the leap from “reduces colitis in mice” to “improves energy metabolism in people” is exactly the kind of extrapolation this article exists to flag.
The second route — KPV’s α-MSH parentage — is where careful readers must be most skeptical, because it is the source of the most seductive and least justified claims. α-MSH is indeed a metabolically important hormone: acting through melanocortin receptors, it is a central regulator of energy homeostasis, and the melanocortin-4 receptor is a critical coordinator of body weight and energy balance.12 α-MSH also has peripheral metabolic actions; for example, it can promote skeletal-muscle glucose uptake and thermogenesis via melanocortin-5 receptors.11 One can see the temptation: KPV is a piece of α-MSH, α-MSH regulates metabolism, therefore KPV must too. This reasoning is a category error. KPV’s defining pharmacological feature is precisely that it does not activate melanocortin receptors — it is the fragment of α-MSH that retains anti-inflammatory activity while being uncoupled from the receptor-mediated melanocortin signaling through which α-MSH exerts its metabolic and pigmentary effects. In other words, KPV was, in effect, stripped of the very mechanism that makes α-MSH metabolically active. Attributing α-MSH’s energy-balance effects to KPV inverts the actual pharmacology.
So KPV’s honest metabolic profile is: a plausible, indirect benefit via inflammation reduction in inflamed states, and essentially no basis for a direct melanocortin-style metabolic effect, because it deliberately bypasses that pathway. Both facts point the same way — toward treating KPV’s metabolic role as an untested hypothesis at best.
What the Evidence Actually Shows — and the Human-Trial Gap
Having examined each component, we can now state the collective evidence position plainly. There are no clinical trials of the KLOW blend for any endpoint, and none for energy metabolism specifically. There are no controlled studies of the four-peptide combination in animals for metabolic outcomes. The individual peptides have their own literatures — substantial for GHK-Cu and BPC-157, moderate for thymosin β4/TB-500, and focused-but-real for KPV — but these literatures concern tissue repair, angiogenesis, gene modulation, and inflammation, not energy metabolism as an endpoint. Where a metabolic link exists, it is inferential (copper as a respiratory cofactor; angiogenesis as a determinant of fuel delivery; inflammation reduction as relief of a metabolic tax), and none of these inferences has been closed with direct data on the blend.
The clearest way to hold this together is a component-by-component evidence table.
| Component | Best-documented biology | Link to energy metabolism | Direct metabolic evidence |
|---|---|---|---|
| GHK-Cu (50 mg) | Gene modulation, tissue remodeling, wound repair, antioxidant support12 | Carries copper, an obligatory cofactor of cytochrome c oxidase (Complex IV)10 | Indirect/cofactor-level only; no human ATP or metabolic-rate data |
| BPC-157 (10 mg) | Angiogenesis via VEGFR2–Akt–eNOS; NO-system modulation345 | Perfusion determines oxygen/fuel delivery to tissue | None; angiogenic effects are preclinical, local, and repair-focused |
| TB-500 (10 mg) | Actin regulation, cell migration, repair, cardioprotection67 | Very tenuous; repair is energetically costly, not energy-producing | None |
| KPV (10 mg) | NF-κB inhibition; anti-inflammatory in colitis models89 | Chronic inflammation imposes a metabolic tax; α-MSH parent is metabolic1112 | None for metabolism; deliberately bypasses melanocortin receptors |
| KLOW blend | Marketed for tissue-repair research1 | Sum of four indirect, unproven links | No trials of any kind |
Two honest conclusions follow. First, the “studies reveal” framing of the title cannot be satisfied at the level of the blend: no study has revealed anything about KLOW and energy metabolism because no such study exists. Second, the component-level science reveals plausible mechanistic threads — a real copper–mitochondria connection, a real perfusion–delivery connection, a real inflammation–metabolism connection — but every one of them is indirect, permissive, and context-dependent, and each has been demonstrated (where at all) for a single peptide against a non-metabolic endpoint. The correct scientific posture toward “KLOW improves energy metabolism” is therefore not “weak evidence” but “untested hypothesis built from adjacent biology.”
The human-trial gap deserves emphasis because it is so often papered over. None of these peptides, individually or combined, is an approved therapy; the human data that exist are sparse and, for BPC-157 and TB-500, essentially absent from rigorous controlled trials. Building a metabolic claim on top of that foundation — and then compounding it by combining four such peptides into a blend never tested as a unit — stacks inference upon inference. For readers who want to see how the sibling GLOW formulation’s repair claims are evaluated, the analysis of whether research supports GLOW in collagen-synthesis pathways models the same measured, evidence-first approach applied to KLOW here.
How KLOW Compares With Compounds Actually Studied for Energy Metabolism
One of the most clarifying exercises is to place KLOW beside compounds that have genuinely been investigated for energy-metabolism endpoints. The contrast is not a competition — KLOW has never entered the metabolic arena — but it shows what a real metabolic candidate looks like in terms of mechanism and evidence, and by that yardstick KLOW’s claim is exposed as aspirational.
| Compound / class | Primary metabolic mechanism | Evidence level for metabolic effect |
|---|---|---|
| Semaglutide / tirzepatide (incretin agonists) | GLP-1 (± GIP) receptor agonism; appetite, insulin, energy balance | Large randomized human trials with approved metabolic indications (type 2 diabetes, obesity) |
| Thyroid hormone (T3/T4) | Sets basal metabolic rate via nuclear receptors | Foundational human endocrinology; approved for hypothyroidism |
| α-MSH / melanocortin agonists | Central and peripheral melanocortin signaling; energy expenditure, glucose uptake1112 | Extensive mechanistic and some clinical data (e.g. MC4R-pathway agonists) |
| Mitochondrial cofactors (e.g. copper, CoQ10, B-vitamins) | Enzyme cofactors for respiration10 | Clear benefit in deficiency; little effect when replete |
| KLOW blend (GHK-Cu/BPC-157/TB-500/KPV) | Tissue repair, angiogenesis, anti-inflammation; only indirect metabolic links | No studies on energy metabolism; investigational, not approved |
The pattern is unmistakable. Genuine metabolic agents act directly on the systems that set energy balance — incretin signaling, thyroid axis, melanocortin receptors, or respiratory cofactor status — and they carry evidence proportionate to their claims, up to and including large approved human trials for the incretin drugs. KLOW acts on repair and inflammation and reaches metabolism only through indirect, unproven bridges. Notably, the one component with an α-MSH lineage (KPV) is precisely the one engineered to lack melanocortin-receptor activity, so it cannot borrow the metabolic credentials of its parent hormone.1112
There is a further lesson embedded in the comparison. Even the mitochondrial-cofactor row — the closest analogue to GHK-Cu’s copper story — comes with a crucial qualifier: cofactors help when you are deficient and do little when you are replete. That single qualifier undercuts most “metabolism-boosting” marketing, because it means the copper in GHK-Cu would only plausibly aid energy metabolism in a copper-limited context, which is not the situation of a typical, adequately nourished person. None of this makes KLOW uninteresting; it makes it a repair-and-regeneration research tool that has wandered, via marketing, into a metabolic conversation it was never equipped to join.
Research Handling and Reconstitution Context
Because KLOW is supplied as a lyophilized (freeze-dried) multi-peptide powder, a brief, strictly educational note on laboratory handling is warranted — with the emphasis that this describes standard research-peptide practice, not a usage recommendation, and that KLOW is not an approved therapeutic for any indication, metabolic or otherwise.
Lyophilized peptides are generally reconstituted with sterile or bacteriostatic water for laboratory purposes. The diluent is directed slowly against the inside wall of the vial rather than sprayed onto the powder, and the vial is swirled gently rather than shaken, because vigorous agitation can shear peptide bonds and denature material — a concern amplified in a four-peptide blend, where four molecules of differing stability must survive the same handling. The chosen diluent volume simply sets the concentration: a fixed mass of blended peptide dissolved in a larger volume yields a lower concentration per unit volume, the arithmetic underlying any reconstitution chart. General educational walkthroughs of this math appear in the site’s peptide reconstitution guide, which is illustrative of how such calculations are typically presented for single peptides and, with more caution, for blends.
Blends introduce a specific complication worth naming: because the four peptides are present at a fixed ratio (here 50:10:10:10 for GHK-Cu:BPC-157:TB-500:KPV), a researcher cannot independently vary one component’s concentration without changing the others. This is a methodological limitation for anyone trying to attribute an observed effect to a particular ingredient, and it is one more reason blends are poorly suited to answering mechanistic questions — including the energy-metabolism question — that would require isolating individual peptides.
| Parameter | Typical research-context practice |
|---|---|
| Lyophilized storage | Cool, dark conditions; long-term stability favored by freezing |
| After reconstitution | Refrigerated; used within a limited window |
| Light and heat | Minimize exposure; both degrade peptides |
| Agitation | Swirl gently; avoid shaking or foaming |
| Freeze-thaw | Repeated cycles degrade peptides; avoid |
| Copper-containing GHK-Cu | Characteristic blue tint; protect from prolonged light; copper adds oxidative-handling considerations |
It bears repeating that meticulous handling changes nothing about the evidence question. A perfectly reconstituted, high-purity vial of KLOW is still a blend with zero energy-metabolism data. Good technique preserves whatever biological activity the molecules have; it does not manufacture efficacy where none has been demonstrated. Researchers cataloguing handling parameters can reference the site’s central dosages index, which is organized for educational comparison rather than as guidance for human use.
Safety, Tolerability, and Combination-Specific Risks
Safety discussion for KLOW must begin with a candid limitation: because the blend has never been formally studied, there is no clinical safety dataset for the combination. What exists is fragmentary safety information on the individual peptides, mostly from preclinical work, plus the general risk profile of unregulated research-chemical products. “No reported major toxicity in rodent studies” is not the same as “safe for human use,” and nothing in this section should be read as an endorsement of use.
Several considerations are specific to a four-peptide blend of this composition:
- Copper load. GHK-Cu is the dominant component by mass (50 of 80 mg), and it delivers copper. While copper is an essential nutrient, excess copper is pro-oxidant and potentially toxic, and repeated administration of a copper-carrying peptide raises theoretical concerns about copper accumulation that have not been characterized for this product. This is one reason the “more is better” intuition is misplaced even for the cofactor argument.
- Angiogenic activity. Both BPC-157 and thymosin β4/TB-500 are pro-angiogenic.37 Promoting new blood-vessel growth is desirable in a healing wound but raises legitimate, if unquantified, caution in any setting where unwanted angiogenesis would be harmful. This is a recognized reason for care with pro-angiogenic peptides and has not been evaluated for the blend.
- Immunomodulation. KPV suppresses NF-κB-driven inflammatory signaling.8 Dampening inflammation is not automatically benign; inflammation is also a protective response, and chronic broad immunosuppression carries its own risks, none of which have been studied for KPV in a blend.
- Product quality. KLOW is sold as a “research chemical” of variable purity outside regulated channels. Impurities, endotoxin, incorrect ratios, and mislabeling are real risks that have nothing to do with the molecules’ intrinsic properties and everything to do with sourcing. In a blend, quality problems in any one of four peptides compromise the whole vial.
- Unknown interactions. The pharmacology of these four peptides administered together — their combined effects, potential interactions, and any additive risks — is simply uncharacterized. Combination products multiply the unknowns.
The reasonable synthesis is that KLOW’s individual components have not thrown up dramatic short-term toxicity signals in the limited preclinical work available, but that this provides no assurance of safety for the untested blend in humans, and certainly no evidence of benefit for energy metabolism or anything else. Absence of demonstrated harm and absence of demonstrated efficacy coexist here, as they do for most research peptides.
Regulatory Status
KLOW’s regulatory position is unambiguous and important to state precisely, because it is frequently blurred in marketing. Neither the KLOW blend nor its individual components is approved by the U.S. Food and Drug Administration, the European Medicines Agency, or any comparable major regulator as a drug for energy metabolism, fatigue, metabolic disease, or any other condition. There is no approved indication of any kind that could be extended to an energy-metabolism use.
The individual peptides sit at various points of regulatory disfavor. BPC-157 has been the subject of explicit FDA caution: it was reviewed in the context of compounding and was not recommended for the Section 503A bulk-drug-substances list, reflecting concern about inadequate characterization and the absence of clinical data. GHK-Cu appears widely in cosmetic formulations, where it is regulated as a cosmetic ingredient rather than as a drug — a status that speaks only to topical cosmetic use, not to systemic administration or metabolic efficacy. TB-500/thymosin β4 and KPV are likewise unapproved for human therapeutic use in these contexts. As a combined injectable blend, KLOW has no recognized regulatory pathway and is sold strictly as a research reagent, typically labeled “for research use only, not for human consumption.”
Two further regulatory points bear on the specific claims this article addresses. First, several of these peptides fall within categories prohibited in sport by the World Anti-Doping Agency — growth-factor and peptide-hormone provisions capture agents like BPC-157 and thymosin β4 — so athletes subject to testing should assume that use of a blend containing them constitutes an anti-doping risk regardless of pharmacology. Second, the cosmetic-ingredient status of GHK-Cu is sometimes cited as though it conferred therapeutic legitimacy; it does not. Approval or acceptance as a topical cosmetic ingredient is a statement about surface-level safety in a cream, not about injected efficacy for metabolism, and conflating the two is one of the more common sleights of hand in peptide-blend marketing.
The regulatory synthesis is straightforward: KLOW is an unapproved, investigational research blend with no recognized therapeutic use, whose components range from cosmetically regulated (GHK-Cu) to explicitly cautioned-against in compounding (BPC-157), and none of which has any approved metabolic indication. Any legitimate investigation of KLOW or its components in energy metabolism would need to proceed through formal preclinical and clinical research under appropriate oversight — not through off-label or informal use.
Frequently Asked Questions
Do studies show that KLOW peptides improve energy metabolism?
No. There are no clinical trials, and no controlled animal studies, of the KLOW blend for energy-metabolism endpoints such as metabolic rate, mitochondrial respiration, glucose disposal, or exercise capacity. The individual peptides have been studied mainly for tissue repair, angiogenesis, gene modulation, and inflammation — not metabolism. Where a metabolic link exists at all, it is indirect and hypothetical: copper as a respiratory cofactor, angiogenesis as a determinant of fuel delivery, and inflammation reduction as relief of a metabolic burden.110 None of these has been demonstrated for the blend in humans.
What is KLOW made of?
KLOW is a fixed blend of four research peptides, most commonly 50 mg GHK-Cu, 10 mg BPC-157, 10 mg TB-500, and 10 mg KPV in an 80 mg lyophilized vial.1 It is essentially the GLOW repair blend (GHK-Cu, BPC-157, TB-500) with the anti-inflammatory tripeptide KPV added. It is marketed as a tissue-repair research reagent, not a metabolic product.
Does the copper in GHK-Cu boost mitochondrial energy production?
Copper is genuinely essential for energy metabolism — it is an obligatory cofactor of cytochrome c oxidase, the final enzyme of the mitochondrial electron transport chain, and copper deficiency impairs ATP production.10 GHK-Cu does carry copper. But there is no evidence that GHK-Cu, at the doses in these blends, measurably increases cytochrome c oxidase activity or ATP output in a living human. Like other cofactors, copper helps mainly when you are deficient and does little when you are replete, and excess copper is potentially harmful. The connection is real biochemistry but an untested, cofactor-level hypothesis for KLOW.
KPV comes from α-MSH, and α-MSH affects metabolism — so does KPV?
This is a common but mistaken inference. α-MSH regulates energy balance through melanocortin receptors, including MC4R centrally and MC5R in muscle.1112 KPV, however, is the α-MSH fragment that retains anti-inflammatory activity while not activating melanocortin receptors — it was, in effect, uncoupled from the very pathway through which α-MSH acts on metabolism. So KPV cannot be assumed to share α-MSH’s metabolic effects; its plausible metabolic relevance runs instead through reducing inflammation, not through melanocortin signaling.
Could reducing inflammation with KPV indirectly help metabolism?
Plausibly, in the right context. Chronic inflammation raises energy expenditure wastefully, worsens insulin resistance, and impairs mitochondrial function, so a compound that dampens NF-κB-driven inflammation could in principle relieve that metabolic tax.8 KPV reduces inflammation in colitis models,89 but it has not been tested for metabolic outcomes, and any such benefit would apply mainly to states of pathological inflammation, not to a healthy person. It remains an untested hypothesis.
Is KLOW approved or safe to use for energy or fatigue?
No. Neither KLOW nor its components is approved by the FDA, EMA, or other major regulators for energy metabolism, fatigue, or any other condition. BPC-157 was specifically not recommended for the FDA’s 503A compounding list; GHK-Cu is regulated as a cosmetic ingredient, which says nothing about injected metabolic efficacy. The blend has no human safety data, carries combination-specific concerns (copper load, dual angiogenic components, immunomodulation, variable product quality), and several components fall under WADA sport prohibitions. It is sold for research use only.
How does KLOW differ from a real metabolic drug like semaglutide or tirzepatide?
Fundamentally. Incretin drugs such as semaglutide and tirzepatide act directly on the GLP-1 (and, for tirzepatide, GIP) receptors that govern appetite, insulin secretion, and energy balance, and they carry large randomized human trials supporting approved metabolic indications. KLOW acts on tissue repair and inflammation, reaches metabolism only through indirect and unproven bridges, and has no trials of any kind. The two are not comparable in mechanism or evidence.
Why do vendors describe KLOW as helping energy or recovery?
Because each ingredient touches biology adjacent to energy — copper and mitochondria, angiogenesis and fuel delivery, inflammation and metabolic cost — and marketing extrapolates from these adjacencies to an energizing claim the data do not support. “Affecting energy metabolism” is also frequently conflated with “giving you energy,” which is a different and unproven proposition. The honest position is that KLOW’s documented biology is regeneration and anti-inflammation, and its metabolic effects are an open, untested question.
How is KLOW handled in a research setting?
As a lyophilized powder, it is reconstituted with sterile or bacteriostatic water using gentle technique (swirl, do not shake), stored cool and dark, and protected from freeze-thaw cycles — standard research-peptide practice, with extra care because it contains four peptides of differing stability plus copper. Because the four components are fixed at a set ratio, researchers cannot vary one without varying the others, which limits mechanistic study. Careful handling preserves activity but does not create metabolic efficacy where none has been demonstrated.
References
- Pickart L, Margolina A. Regenerative and Protective Actions of the GHK-Cu Peptide in the Light of the New Gene Data. Int J Mol Sci. 2018;19(7):1987. PMID: 29986520. https://pubmed.ncbi.nlm.nih.gov/29986520/
- Pickart L, Vasquez-Soltero JM, Margolina A. GHK Peptide as a Natural Modulator of Multiple Cellular Pathways in Skin Regeneration. Biomed Res Int. 2015;2015:648108. PMID: 26236730. https://pubmed.ncbi.nlm.nih.gov/26236730/
- Hsieh MJ, Liu HT, Wang CN, et al. Therapeutic potential of pro-angiogenic BPC157 is associated with VEGFR2 activation and up-regulation. J Mol Med (Berl). 2017;95(3):323-333. PMID: 27847966. https://pubmed.ncbi.nlm.nih.gov/27847966/
- Hsieh MJ, Lee CH, Chueh HY, et al. Modulatory effects of BPC 157 on vasomotor tone and the activation of Src-Caveolin-1-endothelial nitric oxide synthase pathway. Sci Rep. 2020;10:17078. PMID: 33051481. https://pubmed.ncbi.nlm.nih.gov/33051481/
- Sikiric P, Seiwerth S, Skrtic A, et al. Stable Gastric Pentadecapeptide BPC 157 as a Therapy and Safety Key: A Special Beneficial Pleiotropic Effect Controlling and Modulating Angiogenesis and the NO-System. Pharmaceuticals (Basel). 2025;18(6):928. PMID: 40573323. https://pubmed.ncbi.nlm.nih.gov/40573323/
- Goldstein AL, Hannappel E, Sosne G, Kleinman HK. Thymosin β4: a multi-functional regenerative peptide. Basic properties and clinical applications. Expert Opin Biol Ther. 2012;12(1):37-51. PMID: 22074294. https://pubmed.ncbi.nlm.nih.gov/22074294/
- Bock-Marquette I, Saxena A, White MD, DiMaio JM, Srivastava D. Thymosin beta4 activates integrin-linked kinase and promotes cardiac cell migration, survival and cardiac repair. Nature. 2004;432(7016):466-472. PMID: 15565145. https://pubmed.ncbi.nlm.nih.gov/15565145/
- Kannengiesser K, Maaser C, Heidemann J, et al. Melanocortin-derived tripeptide KPV has anti-inflammatory potential in murine models of inflammatory bowel disease. Inflamm Bowel Dis. 2008;14(3):324-331. PMID: 18092346. https://pubmed.ncbi.nlm.nih.gov/18092346/
- Xiao B, Xu Z, Viennois E, et al. Orally Targeted Delivery of Tripeptide KPV via Hyaluronic Acid-Functionalized Nanoparticles Efficiently Alleviates Ulcerative Colitis. Mol Ther. 2017;25(7):1628-1640. PMID: 28143741. https://pubmed.ncbi.nlm.nih.gov/28143741/
- Horn D, Barrientos A. Mitochondrial copper metabolism and delivery to cytochrome c oxidase. IUBMB Life. 2008;60(7):421-429. PMID: 18459161. https://pubmed.ncbi.nlm.nih.gov/18459161/
- Enriori PJ, Chen W, Garcia-Rudaz MC, et al. α-Melanocyte stimulating hormone promotes muscle glucose uptake via melanocortin 5 receptors. Mol Metab. 2016;5(10):807-822. PMID: 27688995. https://pubmed.ncbi.nlm.nih.gov/27688995/
- Krashes MJ, Lowell BB, Garfield AS. Melanocortin-4 receptor-regulated energy homeostasis. Nat Neurosci. 2016;19(2):206-219. PMID: 26814590. https://pubmed.ncbi.nlm.nih.gov/26814590/
Educational and research-use disclaimer: This article is provided solely for scientific and educational purposes. KLOW is an unapproved multi-peptide research blend (GHK-Cu, BPC-157, TB-500, and KPV); neither the blend nor its components is approved by the FDA, EMA, or any comparable regulator for the treatment, cure, or prevention of fatigue, metabolic disease, or any condition of energy metabolism, and no human energy-metabolism efficacy has been demonstrated. The metabolic links discussed here are indirect, mechanistic hypotheses drawn from adjacent biology, not proven effects. Several components fall under WADA sport prohibitions. Nothing here is medical advice or a recommendation for human use. Any legitimate investigation of these compounds should occur within properly authorized preclinical or clinical research under appropriate oversight. Readers should consult qualified professionals and applicable regulations before making any decisions.