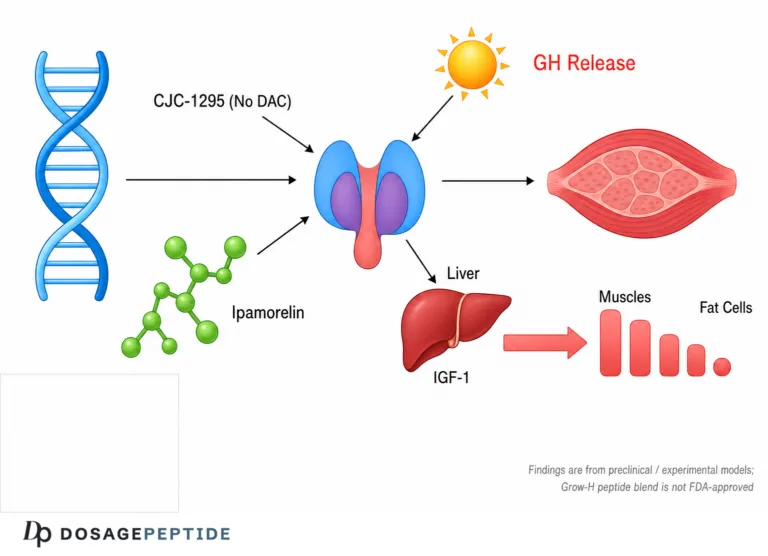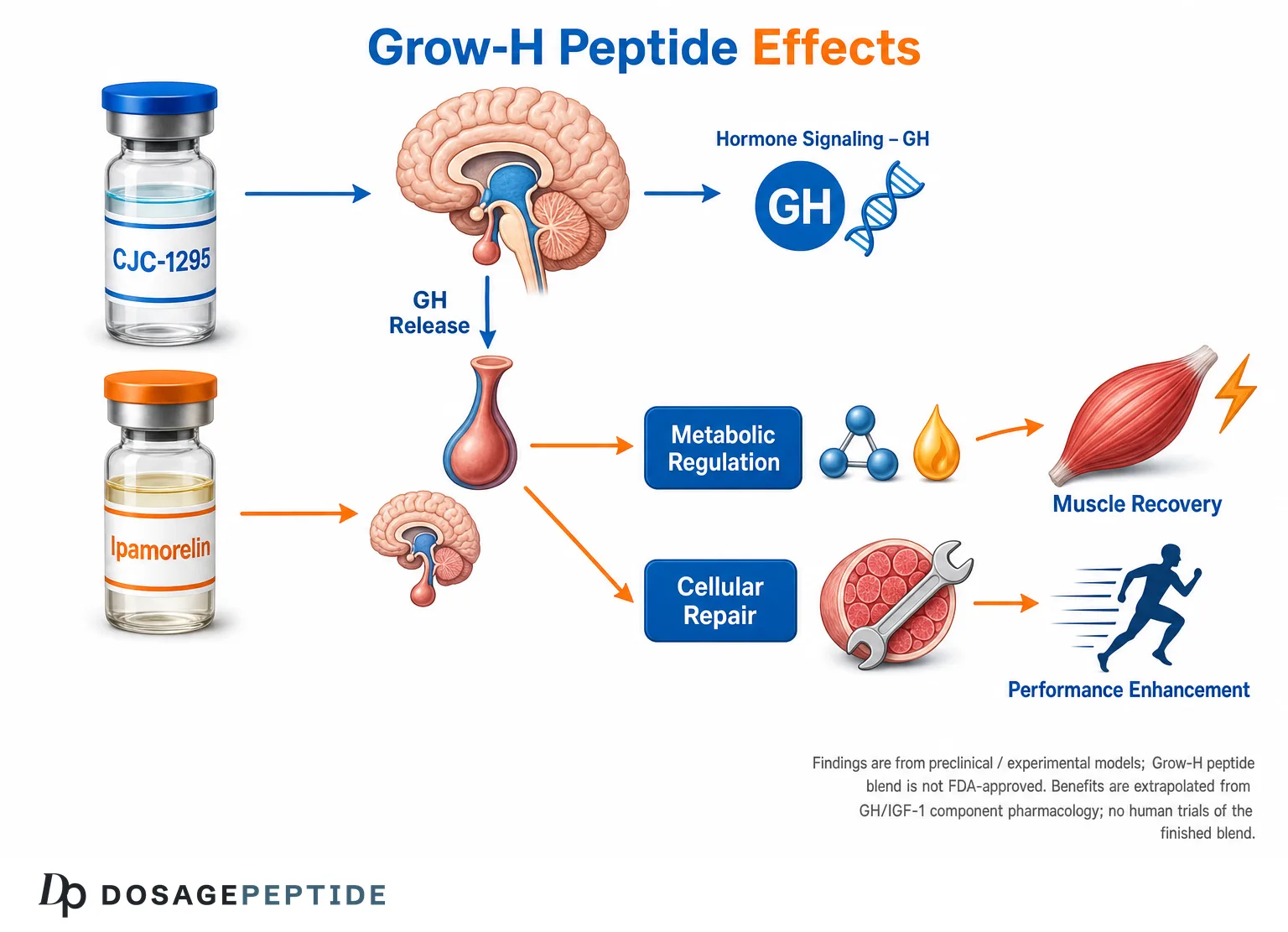
The Grow-H peptide blend is a research-only growth-hormone secretagogue combination — typically pairing a growth-hormone-releasing-hormone (GHRH) analog such as CJC-1295 (no DAC / modified GRF 1–29) with a selective ghrelin-mimetic such as Ipamorelin — that is studied for its ability to amplify the body’s own pulsatile GH output. The central research question behind grow-h peptide blend muscle recovery is whether raising endogenous GH and downstream IGF-1 through this dual-secretagogue approach can meaningfully influence the molecular pathways that govern protein synthesis, tissue repair, and physical performance. The honest short answer, developed in detail below, is that the underlying GH/IGF-1 biology is well characterized, the individual components have real preclinical and early human pharmacology data, but there are no human clinical trials of the finished “Grow-H” blend for muscle recovery or performance — so any recovery claim is an extrapolation, not a proven outcome.
This article is written for educational and research purposes only. Grow-H is not an FDA-approved drug, its constituent peptides are not approved for muscle recovery or performance enhancement in healthy people, and nothing here is medical or dosing advice. The goal is to explain the pharmacology precisely, review the evidence strictly by study type, and mark clearly where confident biology ends and speculation begins.
What Is the Grow-H Peptide Blend?
“Grow-H” is a commercial/research label rather than a single defined molecule. In practice it refers to a fixed-ratio blend of two categories of growth-hormone secretagogue that act on distinct receptors on the pituitary somatotroph and are combined precisely because their mechanisms complement each other. The most common formulation pairs a long-acting or modified GHRH analog with a GHRP-class ghrelin mimetic, and one widely referenced version is built around the CJC-1295 (no DAC) + Ipamorelin 10 mg blend dosage protocol. Understanding Grow-H therefore means understanding two peptides and the logic of stacking them.
A secretagogue is a compound that stimulates secretion of something — here, growth hormone from the anterior pituitary. This is a fundamentally different strategy from injecting recombinant human GH (rhGH) directly. Rather than flooding the circulation with exogenous hormone, secretagogues coax the pituitary to release its own stored GH in a pattern that more closely resembles natural physiology, which is one of the theoretical arguments researchers give for the approach. It also means the effect is constrained by the pituitary’s own capacity and by negative-feedback loops (somatostatin, IGF-1), an important safety and mechanism distinction discussed later.
The GHRH-analog component (CJC-1295 / modified GRF 1–29)
GHRH is the hypothalamic peptide that normally travels down the hypophyseal portal system to tell somatotrophs to synthesize and release GH. Native GHRH is fragile, with a plasma half-life measured in minutes because the enzyme dipeptidyl peptidase-4 (DPP-4) rapidly clips its N-terminus. The bioactive core of GHRH lives in its first 29 amino acids (GRF 1–29, also called sermorelin). CJC-1295 is an engineered GRF 1–29 with four amino-acid substitutions that resist DPP-4 cleavage and other degradation, substantially extending its functional life compared with native GHRH.
There are two distinct molecules commonly discussed under the CJC-1295 name, and conflating them is a frequent source of confusion. CJC-1295 with DAC (Drug Affinity Complex) adds a maleimidopropionyl group that binds covalently to circulating albumin, dramatically prolonging half-life to roughly 5.8–8.1 days and elevating GH and IGF-1 for days after a single dose, as documented in the human pharmacokinetic study by Teichman and colleagues.[2] CJC-1295 without DAC — the version most often used in muscle-recovery-oriented blends and frequently just called “modified GRF 1–29” — lacks the albumin anchor, so it has a short half-life (on the order of ~30 minutes) and produces a sharp, transient bump in GH release rather than a multi-day plateau. Grow-H blends generally use the no-DAC form so that the GHRH signal is pulsatile and can be timed, rather than continuously elevated.
The ghrelin-mimetic / GHRP component (Ipamorelin)
The second arm of the blend acts on an entirely different receptor: the growth-hormone-secretagogue receptor type 1a (GHS-R1a), the same receptor targeted by the stomach hormone ghrelin. GHRPs (growth-hormone-releasing peptides) and non-peptide ghrelin mimetics bind GHS-R1a to trigger GH release and, in the case of many older GHRPs, also to stimulate appetite, cortisol, and prolactin. Ipamorelin is notable because it was characterized as the first selective GH secretagogue: in the foundational work by Raun and colleagues, ipamorelin released GH with a potency and specificity comparable to GHRH but, unlike GHRP-6 and GHRP-2, did not meaningfully raise ACTH/cortisol or prolactin at GH-releasing doses.[4] That selectivity is the main reason Ipamorelin, rather than a more “dirty” GHRP, is the ghrelin-mimetic of choice in modern recovery blends. Readers wanting a component-level primer can review what Ipamorelin is and how it works, and the single-peptide reference point in the Ipamorelin 5 mg dosage protocol.
Why combine two secretagogues instead of one?
The rationale for stacking a GHRH analog with a GHRP is mechanistic, not marketing. The two receptors sit on the same somatotroph but drive different intracellular cascades and interact with different brakes. A GHRH analog raises the amplitude of the GH pulse and increases the pool of GH available for release; a GHRP amplifies release, blunts somatostatin (the inhibitory hypothalamic signal), and recruits somatotrophs that GHRH alone leaves quiescent. When given together, the observed GH output is characteristically greater than the arithmetic sum of each given alone — a synergy repeatedly described in GH-secretagogue physiology. The practical consequence is that a smaller quantity of each peptide can produce a robust, short-lived GH pulse, which is the pattern the Grow-H concept is built around. It is worth stating plainly that this synergy is documented for the components and for GHRH+GHRP co-administration generally; it is the reasonable mechanistic basis for the blend, but it is not the same as a trial proving the branded blend improves recovery.
How Does the Grow-H Blend Trigger Growth Hormone Release?

To understand any downstream recovery claim, you have to start at the somatotroph membrane. The two components of Grow-H converge on GH release through parallel, complementary signal-transduction pathways.
GHRH-receptor signaling: the cAMP/PKA amplitude pathway
The GHRH receptor (GHRHR) is a class B G-protein-coupled receptor coupled to Gs. When the GHRH analog binds, Gs activates adenylate cyclase, raising intracellular cyclic AMP (cAMP). cAMP activates protein kinase A (PKA), which phosphorylates the transcription factor CREB and promotes transcription of the GH1 gene and of Pit-1 (POU1F1), the master transcription factor for somatotroph identity. The net effect is twofold: an acute release of stored GH and a slower increase in GH synthesis, replenishing the releasable pool. This is why the GHRH arm is often described as setting pulse amplitude and supporting the somatotroph’s productive capacity over time.
Ghrelin-receptor signaling: the IP3/Ca2+ release-and-disinhibition pathway
GHS-R1a is a class A GPCR coupled predominantly to Gq/11. Ipamorelin binding activates phospholipase C (PLC), which generates inositol trisphosphate (IP3) and diacylglycerol (DAG). IP3 mobilizes calcium from intracellular stores, and the rise in cytosolic Ca2+ is the proximal trigger for exocytosis of GH-containing secretory granules. Beyond this direct secretory push, GHS-R1a activation acts at the hypothalamic level to suppress somatostatin tone. Because somatostatin is the dominant inhibitory brake on GH release, lifting it allows the GHRH signal to express itself more fully. The ghrelin arm therefore contributes both a direct calcium-driven release and a disinhibition that widens the window for GHRH to act.
Convergence at the somatotroph: amplitude, frequency, and pulse fidelity
The synergy of the blend is best understood as complementary control of a pulse. GHRH raises how much GH each pulse contains and keeps the factory stocked; the ghrelin mimetic increases the probability and size of release while removing the somatostatin brake. Combined, they produce a larger, cleaner GH pulse that then decays — preserving the pulsatile shape that peripheral tissues appear to “read” differently from continuous exposure. This pulsatility argument is a genuine theoretical advantage of secretagogues over continuous rhGH, though whether it translates into superior tissue outcomes in humans is unproven.
| Feature | GHRH analog arm (CJC-1295 / mod GRF 1–29) | Ghrelin-mimetic arm (Ipamorelin) |
|---|---|---|
| Target receptor | GHRH receptor (GHRHR), class B GPCR | GHS-R1a (ghrelin receptor), class A GPCR |
| Primary G-protein | Gs | Gq/11 |
| Second messenger | ↑ cAMP → PKA → CREB / Pit-1 | ↑ IP3/DAG → intracellular Ca2+ release |
| Chief effect on GH pulse | Raises pulse amplitude; supports GH synthesis | Triggers release; suppresses somatostatin (disinhibition) |
| Effect on appetite | Minimal | Modest (GHS-R1a is the ghrelin receptor); less than GHRP-6 |
| Cortisol / prolactin | No meaningful rise | Selective — little to no rise at GH-releasing doses[4] |
| Feedback still intact? | Yes — IGF-1 and somatostatin negative feedback preserved | Yes — pituitary-capped, feedback-regulated |
A Closer Look at Receptor Biology: Constitutive Activity, Signaling Bias, and Desensitization
The somatotroph-level summary above treats the two receptors as simple on-switches, but the deeper pharmacology is more interesting and has direct implications for how a pulsatile blend is designed. Two features stand out: the ghrelin receptor’s unusually high baseline activity, and the way both receptors adapt to repeated stimulation.
The ghrelin receptor is constitutively active
Unlike most GPCRs, GHS-R1a is not quiescent in the absence of ligand. In classic transfected-cell work, the ghrelin receptor signaled at roughly 50% of its maximal capacity with no agonist present at all — one of the highest ligand-independent (constitutive) activities described for any GPCR — and the same study identified a potent inverse agonist able to switch that basal tone off.[9] This matters for interpretation: a fraction of GHS-R1a signaling proceeds regardless of how much Ipamorelin is present, and the receptor’s basal tone is thought to contribute to physiological set-points such as GH pulsatility and appetite drive. It also complicates any simple dose-response picture, because the agonist is modulating a system that already carries a standing signal rather than switching a silent receptor on from zero.
Signaling beyond the classic second messengers
Both receptors do more than raise a single second messenger. GHS-R1a, while predominantly Gq/11-coupled, can also engage Gi/o and recruit β-arrestin, and β-arrestin recruitment governs receptor internalization and can drive its own (G-protein-independent) signaling. GHRHR signaling likewise is not only about acute cAMP: the CREB/Pit-1 arm feeds back on somatotroph proliferation and GH-gene transcription over longer timescales. The practical point is that which pathway a ligand favors — so-called biased agonism — can in principle differentiate secretagogues that look identical on a simple GH-release readout. That nuance is characterized for some ghrelin-receptor ligands in general pharmacology but has not been mapped in detail for the specific Grow-H components, so it should be treated as a mechanistic possibility rather than an established feature of the blend.
Desensitization, internalization, and why pulses beat plateaus
Sustained agonist exposure at a GPCR typically triggers receptor phosphorylation, β-arrestin binding, internalization, and downregulation — the receptor becomes less responsive over time (tachyphylaxis). For GH secretagogues this is not a theoretical footnote: continuous GHRH or continuous ghrelin-receptor stimulation tends to blunt the GH response, whereas intermittent, pulsatile stimulation better preserves responsiveness. This receptor-level logic is a second, independent argument — alongside the “tissues read pulses” argument — for favoring short-acting components dosed as discrete pulses rather than a molecule that holds the receptor occupied for days. It is also part of why the no-DAC CJC-1295 form, rather than the multi-day DAC form, is generally chosen for a recovery-oriented blend.
From a GH Pulse to Muscle Tissue: The GH/IGF-1 Axis and Recovery Pathways
A GH pulse is only the first domino. The recovery-relevant biology unfolds downstream, through both direct GH actions on tissues and the GH-stimulated production of insulin-like growth factor 1 (IGF-1). This axis — its physiology reviewed comprehensively in the endocrine literature — is where the plausible connection to muscle repair lives.[1]
Systemic IGF-1 versus local mechano-growth factor
When GH reaches the liver, it binds the GH receptor and, via JAK2/STAT5 signaling, drives hepatic transcription of IGF-1. This liver-derived IGF-1 circulates (largely bound to IGFBP-3 and the acid-labile subunit) and accounts for most of the IGF-1 measured in blood. But skeletal muscle also produces IGF-1 locally, and mechanical loading favors a splice variant sometimes called mechano-growth factor (MGF, IGF-1Ec). The distinction matters: circulating IGF-1 provides a permissive anabolic background, whereas locally expressed IGF-1 acts in an autocrine/paracrine fashion right where mechanical damage and remodeling are occurring. Secretagogues raise systemic GH and therefore systemic IGF-1; they do not by themselves supply the mechanical stimulus that drives the local, load-specific IGF-1 response — which is one reason exercise is treated as the indispensable partner variable in any serious discussion of muscle outcomes.
PI3K/Akt/mTOR: the protein-synthesis engine
IGF-1 (and insulin) binding to the type 1 IGF receptor activates the PI3K/Akt pathway, and Akt sits at a hub with outsized influence over muscle mass. Akt activates mTORC1, which upregulates protein translation through S6K1 and 4E-BP1, increasing the rate of muscle-protein synthesis. Simultaneously, Akt phosphorylates and inactivates the FoxO transcription factors, which suppresses the muscle-specific ubiquitin ligases atrogin-1 (MAFbx) and MuRF1 — the machinery of protein breakdown. In other words, IGF-1 signaling can shift net protein balance in the anabolic direction from both sides: more synthesis, less degradation. This PI3K/Akt axis, activated by both exercise and IGF-1/insulin, is the widely cited central regulator of muscle mass in the pharmacology literature.[5]
Satellite cells and myonuclear addition
Beyond acute protein turnover, meaningful hypertrophy and repair depend on satellite cells — the resident muscle stem cells that, upon injury or overload, proliferate, differentiate, and fuse with existing fibers to donate new myonuclei. IGF-1 is a well-established driver of satellite-cell proliferation and of their fusion into fibers, a mechanism detailed in reviews of muscle-mass regulation.[5] This is the theoretical bridge from “more IGF-1” to “better repair after a damaging bout,” because supplying more myonuclei expands the transcriptional capacity of a fiber to rebuild. It is a plausible pathway — and it is exactly the kind of mechanism that has been demonstrated with IGF-1 in cell and animal models but not validated as a recovery outcome for the Grow-H blend in humans.
Direct GH actions: lipolysis, nitrogen retention, and connective tissue
Not all of GH’s relevant effects run through IGF-1. GH acts directly on adipocytes to promote lipolysis, mobilizing free fatty acids for fuel, and it exerts a nitrogen-retaining, protein-sparing effect — particularly evident during stress and fasting, where GH functions as a lean-mass-preserving anabolic/anti-catabolic signal. The human metabolic-effects literature describes GH as a primary anabolic hormone in these states, stimulating fat oxidation while conserving lean body mass and inducing a degree of insulin resistance.[3] GH and IGF-1 also influence connective tissue: they stimulate collagen synthesis in tendon and other extracellular-matrix compartments, a mechanism often invoked to explain why GH-axis manipulation is studied in the context of tendon, ligament, and joint recovery rather than muscle alone. These direct actions broaden the plausible recovery footprint but do not, on their own, equal proven performance benefit.
| Signaling step / effector | Trigger | Recovery-relevant consequence |
|---|---|---|
| Hepatic GHR → JAK2/STAT5 | GH pulse reaching liver | ↑ circulating IGF-1 (permissive anabolic background) |
| Local muscle IGF-1 / MGF | Mechanical load + GH milieu | Autocrine/paracrine repair signal at damaged fibers |
| IGF-1R → PI3K/Akt → mTORC1 | IGF-1 / insulin | ↑ muscle-protein synthesis (S6K1, 4E-BP1) |
| Akt → FoxO inhibition | IGF-1 signaling | ↓ atrogin-1 / MuRF1 → less protein breakdown |
| IGF-1 → satellite cells | IGF-1 + injury/overload | Proliferation, fusion, new myonuclei for repair |
| GH → adipocyte lipolysis | GH pulse | Fatty-acid mobilization; body-recomposition signal |
| GH/IGF-1 → collagen synthesis | GH pulse | Tendon/ECM remodeling; connective-tissue repair |
What Does the Evidence Actually Show, Organized by Study Type?
This is the section that matters most for an honest YMYL treatment. The evidence for Grow-H is emphatically component-level and spans in-vitro, animal, and early human-pharmacology work — with a hard ceiling at “GH/IGF-1 secretion” endpoints rather than validated muscle-recovery or performance outcomes. There is no clinical trial of the assembled “Grow-H” product. For a curated look at the underlying literature, see the evidence supporting the Grow-H peptide blend.
In-vitro and cellular evidence
At the cell level, the pieces of the mechanism are well supported. Somatotroph and pituitary cell models established that GHRH acts through cAMP/PKA and that GHRPs/ghrelin mimetics act through GHS-R1a and calcium mobilization; the receptor pharmacology of ipamorelin (selective GHS-R1a agonism releasing GH without cortisol/prolactin activation) was characterized in exactly this kind of work.[4] On the muscle side, cell-culture systems (myoblasts, myotubes) have repeatedly shown that IGF-1 stimulates myotube protein synthesis, satellite-cell proliferation, and hypertrophy via PI3K/Akt/mTOR, forming the mechanistic backbone reviewed in muscle-mass regulation.[5] What in-vitro data cannot do is prove that a secretagogue-driven GH pulse produces enough local IGF-1 signaling, in a whole organism, to translate into faster real-world recovery. Cell dishes lack the endocrine feedback, pharmacokinetics, and mechanical loading of a living muscle.
Animal evidence
Animal models establish that the GHRH-analog approach actually works in a whole organism. In the GHRH-knockout mouse, once-daily CJC-1295 restored normal growth, demonstrating that a stabilized GRF 1–29 analog can drive the GH/IGF-1 axis in vivo and rescue a GH-deficient phenotype.[8] Ipamorelin’s original characterization also included animal work showing dose-dependent GH release and, notably, effects on longitudinal bone growth and body-weight gain in rodents, consistent with a functioning GH/IGF-1 output.[4] Separately, direct IGF-1 delivery to rodent muscle produces genuine hypertrophy, and local IGF-1 overexpression models support the satellite-cell mechanism. The caveats are the usual ones: rodents are not humans, GH physiology and IGF-1 responsiveness differ across species, and “normalizes growth in a deficient animal” is not the same claim as “accelerates recovery in a trained, healthy human.”
Human pharmacology evidence (component level)
The strongest human data are pharmacologic: they show that these compounds raise GH and IGF-1 in people, which is the necessary first link in the chain. The pivotal human study of CJC-1295 (with DAC) in healthy adults documented dose-dependent increases in GH (2- to 10-fold) sustained for six days or more and IGF-1 (1.5- to 3-fold) elevated for 9–11 days after a single injection, with a half-life of roughly 5.8–8.1 days and cumulative IGF-1 elevation over repeated dosing.[2] That establishes proof-of-pharmacology for the GHRH arm in humans. For the ghrelin arm, human and translational data support that selective secretagogues raise GH/IGF-1; the closest long-term human read on a ghrelin-mimetic secretagogue comes from the oral compound MK-677 (ibutamoren), where a two-year randomized trial in older adults increased IGF-1 into the young-adult range and increased fat-free mass by roughly 1.1 kg — but did not improve strength and worsened insulin sensitivity.[7] That last study is a crucial honesty anchor: raising IGF-1 and even lean mass does not automatically deliver the functional outcome (strength) that most people actually care about.
The critical gap: no human trial of the finished Grow-H blend
Putting it bluntly: as of this writing there is no published randomized controlled trial — indeed no published clinical trial of any kind — testing the assembled “Grow-H” CJC-1295-plus-Ipamorelin blend for muscle recovery, hypertrophy, or athletic performance in humans. Every performance or recovery claim is a chain of inferences: components raise GH → GH raises IGF-1 → IGF-1 activates known anabolic pathways → those pathways are involved in repair. Each individual link has support, but the assembled clinical claim does not. A candid discussion of this gap, and of what would be needed to close it, is covered in clinical insights on whether the Grow-H blend boosts strength and repair.
| Study type | What is genuinely shown | What it does not show | Evidence strength |
|---|---|---|---|
| In-vitro / cellular | Receptor pharmacology (GHRHR cAMP; GHS-R1a Ca2+); IGF-1 drives myotube synthesis & satellite-cell proliferation | Whole-organism translation; recovery outcomes | Strong for mechanism, none for outcome |
| Animal | CJC-1295 rescues growth in GHRH-KO mice; ipamorelin raises GH/bone growth; local IGF-1 causes hypertrophy | Human relevance; trained-athlete recovery | Moderate, mechanism-confirming |
| Human pharmacology (components) | CJC-1295 raises GH/IGF-1 for days; secretagogues raise IGF-1; MK-677 raises IGF-1 & lean mass | Strength gains; recovery endpoints; the blend specifically | Moderate for GH/IGF-1 endpoints only |
| Human clinical (the Grow-H blend) | — nothing published — | All muscle-recovery/performance claims | Absent |
The Broader GHRP and GHRH-Analog Family: Where Ipamorelin and CJC-1295 Sit
Ipamorelin and CJC-1295 did not appear in isolation; each belongs to a well-characterized family of GH secretagogues developed over decades of endocrine research. Understanding the neighbors clarifies exactly what the Grow-H components were selected to do — and to avoid. The GHRP lineage traces back to the discovery that certain synthetic enkephalin-derived peptides release GH through a receptor distinct from the GHRH receptor — the pathway later identified as GHS-R1a.[10]
The GHRP / ghrelin-mimetic side of the family
GHRP-6 was the prototype and is the most strongly appetite-stimulating; it also raises cortisol and prolactin more than newer agents. GHRP-2 (pralmorelin) is a potent GH releaser used diagnostically to probe pituitary GH reserve, but it likewise carries some cortisol/prolactin activity. Hexarelin is the most potent GHS-R1a agonist of the classic trio and has a distinctive additional interaction with the CD36 scavenger receptor implicated in cardiovascular tissue, yet it is comparatively prone to receptor desensitization with continued use. Ipamorelin was engineered later specifically to keep GHRH-comparable GH-releasing potency while shedding the off-target cortisol/prolactin activation — the selectivity documented in its original characterization[4] — which is precisely why it, rather than a “stronger” but dirtier GHRP, is the ghrelin arm in a modern blend. MK-677 (ibutamoren) sits in the same functional class but is an orally active non-peptide with long duration, giving tonic rather than pulsatile IGF-1 elevation.
The GHRH-analog side of the family
On the GHRH side, sermorelin is simply GRF 1–29 — the shortest fully active fragment — and is the least modified and shortest-acting member. Tesamorelin is a stabilized GHRH analog that is the one FDA-approved member of this family, but only for HIV-associated lipodystrophy, not for performance or recovery. CJC-1295 no-DAC (modified GRF 1–29) adds degradation-resistant substitutions to GRF 1–29 for a slightly longer but still pulsatile action, while CJC-1295 with DAC adds the albumin-binding complex for multi-day duration. The Grow-H concept deliberately draws from the “short-acting, pulsatile, selective” end of both families.
| Compound | Class | Receptor | Notable feature | Typical duration / pattern |
|---|---|---|---|---|
| GHRP-6 | Peptide GHRP | GHS-R1a | Strong appetite; higher cortisol/prolactin | Short pulse |
| GHRP-2 (pralmorelin) | Peptide GHRP | GHS-R1a | Potent releaser; diagnostic use; some cortisol/prolactin | Short pulse |
| Hexarelin | Peptide GHRP | GHS-R1a (+ CD36) | Most potent of trio; desensitization-prone | Short pulse |
| Ipamorelin | Peptide GHRP | GHS-R1a | Selective — minimal cortisol/prolactin[4] | Short pulse (~2 h) |
| MK-677 (ibutamoren) | Non-peptide mimetic | GHS-R1a | Orally active; long-acting | Tonic IGF-1 elevation |
| Sermorelin | GHRH analog | GHRHR | GRF 1–29; least modified | Very short |
| Tesamorelin | GHRH analog | GHRHR | FDA-approved (HIV lipodystrophy only) | Short–moderate |
| CJC-1295 no DAC | GHRH analog | GHRHR | Stabilized GRF 1–29 | Short pulse (~30 min) |
| CJC-1295 with DAC | GHRH analog | GHRHR | Albumin-bound complex | Multi-day (~5.8–8.1 d)[2] |
Does Grow-H Actually Improve Muscle Recovery and Performance?
The most intellectually honest answer separates three questions that are often blurred together: (1) does the blend raise GH/IGF-1? (2) does raising GH/IGF-1 change body composition? and (3) does it improve functional performance and recovery? The evidence gets weaker as you move down that list.
On question 1, the answer is a qualified yes at the component level: the GHRH arm demonstrably raises GH and IGF-1 in humans,[2] and selective ghrelin mimetics do the same. On question 2, the honest answer is “modestly, mostly fat-free mass and fat mass shifts” — the MK-677 trial showed a ~1.1 kg fat-free-mass gain, and GH itself alters body composition.[7] On question 3, the evidence is genuinely discouraging for the strongest claims. The most rigorous synthesis of GH and athletic performance — a systematic review of 27 study samples and 303 GH-treated participants — concluded that GH increased lean body mass but did not improve strength or exercise capacity, and if anything may have worsened exercise capacity and increased adverse events such as fluid retention and fatigue.[6] Because Grow-H’s entire theory of action routes through the GH/IGF-1 axis, that finding is directly relevant: elevating GH is not a reliable route to strength or performance in already-healthy people, even though it changes body composition.
Where does that leave the recovery question specifically? Recovery is not identical to strength or sprint performance — it encompasses connective-tissue repair, protein re-synthesis after damage, sleep quality, and reduced catabolism. GH’s nitrogen-sparing effect, its collagen-stimulating action on tendon/ECM, and IGF-1’s role in satellite-cell-mediated repair make a plausible case that a secretagogue could support recovery-adjacent processes, and some secretagogues (notably MK-677) improve sleep architecture, which is itself a recovery variable. But “plausible mechanism plus improved sleep” is not a measured recovery outcome for the Grow-H blend. The correct scientific posture is: mechanistically coherent, unproven at the product level, and constrained by the sobering GH-performance literature.
Pharmacokinetics: How the Two Components Behave in the Body
Pharmacokinetics explains why the blend is dosed the way it is and why the no-DAC form is preferred for a pulsatile strategy. The key parameters differ sharply between the GHRH-analog variants and the ghrelin mimetic.
CJC-1295 with DAC is the long-acting molecule: by covalently binding albumin it achieves a multi-day half-life (~5.8–8.1 days) and holds GH/IGF-1 elevated continuously for days after one dose.[2] That continuous elevation is precisely what a pulsatile strategy tries to avoid, because tonic GH exposure departs from natural physiology and blunts the pulse pattern peripheral tissues respond to. CJC-1295 no DAC (modified GRF 1–29) has a short half-life (~30 minutes), so it produces a brief GHRH signal that can be timed and repeated. Ipamorelin similarly has a short half-life (on the order of ~2 hours), giving a transient GH-releasing push. Combining two short-acting agents is what preserves the sharp pulse; the components are typically administered together so their GH-releasing windows overlap. The GH pulse they generate is itself brief (GH’s own circulating half-life is only minutes), but the downstream IGF-1 signal is far more sustained because IGF-1 circulates protein-bound with a half-life of many hours.
| Parameter | CJC-1295 with DAC | CJC-1295 no DAC (mod GRF 1–29) | Ipamorelin |
|---|---|---|---|
| Class | Long-acting GHRH analog | Short-acting GHRH analog | Selective GHRP / ghrelin mimetic |
| Receptor | GHRHR | GHRHR | GHS-R1a |
| Approx. half-life | ~5.8–8.1 days[2] | ~30 minutes | ~2 hours |
| GH/IGF-1 pattern | Sustained, tonic elevation for days | Sharp, transient pulse | Sharp, transient pulse |
| Role in a pulsatile blend | Generally avoided (tonic) | Amplitude / pool support | Release trigger + somatostatin disinhibition |
| Physiologic fidelity | Lower (continuous) | Higher (pulsatile) | Higher (pulsatile, selective) |
Analytical Methods and Assays Used to Study GH Secretagogues
Reading the secretagogue literature critically requires knowing how the numbers are generated, because each assay answers a different question and carries its own limitations. The methods span molecular pharmacology, endocrine physiology, and analytical chemistry, and a claim is only as strong as the assay behind it.
Confirming receptor engagement in vitro
At the receptor level, the workhorses are functional cell assays. cAMP-accumulation assays — reporter genes driven by cAMP-response elements, or direct cAMP quantitation — read out Gs-coupled GHRHR activation, while intracellular-calcium-flux assays (typically fluorescent Ca2+ indicators in cells expressing GHS-R1a) capture the Gq/PLC/IP3 response a ghrelin mimetic triggers. Radioligand or fluorescent competitive-binding assays establish affinity and selectivity, and β-arrestin-recruitment assays probe internalization and bias. These are exactly the kinds of assays that established Ipamorelin’s selective GHS-R1a agonism without cortisol/prolactin-pathway activation,[4] and the same reporter methodology revealed the ghrelin receptor’s high constitutive signaling.[9]
Measuring the hormone output
Moving to the whole organism, the primary endpoint is circulating hormone. Serum GH and IGF-1 are quantified by immunoassays — historically radioimmunoassay (RIA), now more often ELISA/chemiluminescent immunoassay or, for reference-grade work, liquid chromatography–mass spectrometry. Because GH is secreted in sharp pulses and cleared within minutes, a single random GH value is nearly uninterpretable; researchers use frequent timed sampling after a stimulus (a GH-stimulation protocol) and often report peak GH and area-under-the-curve. IGF-1, being protein-bound and stable over hours to days, is the more robust integrated readout of axis activation — which is why the pivotal CJC-1295 human study tracked IGF-1 over many days as its main proof-of-pharmacology.[2] IGFBP-3 and the acid-labile subunit are sometimes measured alongside to interpret IGF-1 bioavailability.
Verifying the material itself
A category often overlooked outside formal labs is analytical confirmation of the peptide. High-performance liquid chromatography (HPLC) assesses purity and detects degradation products, while mass spectrometry confirms molecular identity and correct sequence. For a two-component blend, these methods are the only way to verify that both peptides are present, intact, and in the intended ratio — a real concern given that research-grade materials are not made to pharmaceutical release specifications. Endotoxin and sterility testing address the injectable-solution risks. When a study or a supplier reports “purity,” it almost always means an HPLC number, which speaks to identity and contamination but not to biological potency — two peptides can each be “99% pure” and still be blended in the wrong ratio.
Dose-Response, Biomarkers, and the Ceiling Imposed by Feedback
A recurring misconception is that more secretagogue linearly means more GH, more IGF-1, and more result. The real dose-response is shaped — and capped — by the same feedback biology that makes secretagogues theoretically safer than exogenous GH.
Why the dose-response plateaus
Because a secretagogue works through the pituitary, its output is limited by the releasable GH pool and by negative feedback from rising IGF-1 and somatostatin. Past a certain point, adding more GHRH-analog or ghrelin-mimetic does not produce a proportionally larger GH pulse: the somatotroph has a finite store, and elevated IGF-1 feeds back to restrain further release. This self-limiting behavior is a genuine mechanistic contrast with rhGH, which can push circulating GH to supraphysiologic levels regardless of feedback. It also means the marginal benefit of escalating a secretagogue dose falls off, while the side-effect burden (fluid retention, glucose effects) does not necessarily plateau in the same way — a poor trade at the top of the curve.
Which biomarkers actually track the axis
In research settings, IGF-1 is the biomarker of record because it integrates the pulsatile GH signal into a stable daily value; a sustained rise in IGF-1 (ideally interpreted against age- and sex-specific reference ranges) is the clearest evidence that a secretagogue is engaging the axis. Fasting glucose, HbA1c, and insulin-sensitivity indices are the safety-side biomarkers, given GH’s counter-regulatory, insulin-desensitizing action.[3] Body-composition readouts (DXA fat-free mass and fat mass) sit further downstream and are more meaningful than a single hormone value but still upstream of function. The endpoints that would actually settle the recovery question — muscle protein-synthesis rate, strength, tendon-repair kinetics — remain, as emphasized throughout, uncollected for the blend.
Interpreting biomarker changes honestly
The MK-677 and rhGH literature together deliver the essential cautionary lesson: an impressive biomarker move need not translate into the outcome people care about. MK-677 pushed IGF-1 into the young-adult range and added fat-free mass yet did not improve strength and worsened insulin sensitivity,[7] and the systematic review of GH found lean-mass gains without strength or exercise-capacity improvement.[6] So a favorable IGF-1 shift from a Grow-H-type blend would confirm the pharmacology is working exactly as designed — and would still say nothing definitive about recovery or performance. Biomarkers verify mechanism; they do not certify benefit.
How Does Grow-H Compare to Other GH-Axis Strategies?
Placing the blend against its alternatives clarifies both its theoretical appeal and its limits. The main comparators are exogenous recombinant GH, oral non-peptide secretagogues, and single-agent GHRH analogs.
Compared with recombinant human GH (rhGH), the secretagogue approach is pituitary-dependent and feedback-preserved: because Grow-H asks the pituitary to release its own GH, output remains subject to somatostatin and IGF-1 negative feedback, theoretically limiting supraphysiologic overshoot. rhGH bypasses all of that, delivering hormone regardless of feedback, which is more potent but also removes a natural safety valve. Crucially, though, the performance ceiling identified for rhGH — body-composition change without strength gains, plus adverse effects[6] — is likely to constrain a secretagogue-driven axis too, since both funnel through GH/IGF-1. Compared with MK-677 (ibutamoren), an orally active non-peptide ghrelin mimetic, Grow-H’s injectable pulsatile design contrasts with MK-677’s once-daily oral, more tonic IGF-1 elevation; the two-year MK-677 data usefully preview what sustained IGF-1 elevation does and doesn’t do (lean mass up, strength unchanged, insulin sensitivity worse).[7] Compared with a single GHRH analog alone (e.g., sermorelin/tesamorelin or CJC-1295 by itself), adding the ghrelin arm is what recruits the synergy and somatostatin disinhibition described earlier.
| Strategy | How it raises GH/IGF-1 | Pulsatility | Feedback preserved? | Approval status |
|---|---|---|---|---|
| Grow-H blend (GHRH analog + Ipamorelin) | Dual secretagogue on somatotroph | High (short-acting components) | Yes | Not FDA-approved; research only |
| Recombinant human GH (rhGH) | Direct exogenous GH | Depends on regimen; often tonic | No (bypasses feedback) | Approved for specific medical indications, not healthy performance |
| MK-677 (ibutamoren) | Oral ghrelin mimetic (GHS-R1a) | Lower (sustained IGF-1) | Yes | Investigational; not approved |
| GHRH analog alone (sermorelin/tesamorelin/CJC-1295) | Single GHRH signal | Moderate–high (no-DAC) / low (DAC) | Yes | Tesamorelin approved for HIV lipodystrophy only; others research/legacy |
Research Handling, Reconstitution, and Experimental-Design Context
Because Grow-H is a research material, correct handling is part of getting interpretable results. These peptides are supplied as lyophilized (freeze-dried) powder and must be reconstituted with bacteriostatic water before use in a laboratory setting. Reconstitution technique, concentration math, and storage all affect the actual amount of intact peptide delivered, so precision here is not optional. A full walkthrough of technique is provided in the peptide reconstitution guide, and concentration/volume calculations can be worked out with the dosage calculator.
Several handling variables matter specifically for GH secretagogues. Reconstitution and storage: lyophilized peptide is comparatively stable, but once in solution these peptides degrade with time, heat, light, and agitation; refrigeration and gentle handling (adding diluent to the vial wall rather than jetting it onto the powder) preserve integrity. Timing relative to feeding: in experimental designs, a substantial carbohydrate or fat load near administration can blunt the GH response, because elevated glucose/insulin and free fatty acids suppress GH release; this is why fasting-state administration is a common design control. Timing relative to sleep: the largest natural GH pulse occurs at sleep onset, so pre-sleep administration is frequently modeled to stack the secretagogue pulse on the endogenous nocturnal pulse. Ratio and co-administration: because the synergy depends on both receptors being engaged in the same window, the GHRH-analog and ghrelin-mimetic arms are typically combined so their release windows overlap. For terminology and unit conventions used throughout this literature, the peptide glossary is a useful reference.
On experimental models: the endpoints used to study these compounds ascend a hierarchy of rigor. The most basic is a GH stimulation test — measuring serum GH at intervals after administration to confirm a pulse. Next is IGF-1 response over days to weeks, a more integrated readout of axis activation (this is what the CJC-1295 human study tracked[2]). Body-composition endpoints (DXA-measured fat-free mass and fat mass) are more meaningful but still upstream of function. The endpoints that actually answer the recovery/performance question — strength testing, exercise capacity, biopsy-confirmed protein-synthesis rates, time-to-recovery after a standardized damaging bout — are precisely the ones that have not been collected for the Grow-H blend. Recognizing which endpoint a given claim rests on is the single most useful skill for reading this literature critically.
What Safety Signals Are Reported in the Research?
Because Grow-H itself is unstudied clinically, the safety picture is assembled from component and GH-axis data and should be read as such. Several signals recur.
Fluid retention, joint symptoms, and paresthesia
GH promotes sodium and water retention, and the GH-performance literature specifically flagged fluid retention (edema), joint discomfort/arthralgia, and carpal-tunnel-type symptoms among the more common adverse effects associated with raising GH, along with reduced exercise capacity in some participants.[6] Anything that pushes the GH/IGF-1 axis upward can plausibly reproduce these, and they tend to be dose-related.
Glucose and insulin sensitivity
GH is a counter-regulatory hormone that induces insulin resistance as part of its normal metabolic role.[3] Consistent with this, sustained secretagogue exposure has measurably worsened insulin sensitivity and raised fasting glucose — observed directly in the long-term MK-677 trial.[7] This is arguably the most important metabolic caveat for any GH-axis strategy and a reason continuous/tonic elevation is viewed less favorably than brief pulses.
Selectivity: cortisol, prolactin, and appetite
A point in Ipamorelin’s favor is selectivity: unlike GHRP-6 and GHRP-2, it releases GH without meaningfully raising cortisol or prolactin at GH-releasing doses, which reduces the endocrine “noise” and side-effect burden associated with older GHRPs.[4] Because GHS-R1a is the ghrelin receptor, some appetite stimulation is still expected, though less pronounced than with GHRP-6.
IGF-1 and theoretical proliferative concerns
IGF-1 is mitogenic, and the GH/IGF-1 axis has documented links to tissue growth and, in the broader endocrine literature, to cancer biology — a reason chronic supraphysiologic IGF-1 elevation is treated cautiously.[1] This is a theoretical, long-horizon concern rather than a demonstrated outcome of short-term secretagogue use, but it is a legitimate part of the risk framing, particularly for anyone with a personal or family history of neoplasia. It reinforces why these compounds are research materials, not casual supplements.
Product-quality and unknown-blend risks
A distinct category of risk is not pharmacologic at all: research-grade blends are not manufactured to pharmaceutical standards, so purity, actual peptide content, correct component ratio, and sterility can vary between sources. Because there is no clinical trial of the finished blend, there is also no standardized safety database, no established dosing, and no regulatory oversight of the specific product. That uncertainty is itself a safety consideration and one more reason the honest framing is “research/experimental use only.”
Limitations and Open Questions
Several gaps define the current state of knowledge and would need to be addressed before any confident recovery claim could be made:
- No product-level clinical data. The single largest limitation is the complete absence of human trials on the assembled Grow-H blend for recovery, hypertrophy, or performance. Everything is extrapolated from components and GH/IGF-1 physiology.
- The strength paradox. Raising GH/IGF-1 reliably changes body composition but has repeatedly failed to improve strength or exercise capacity in controlled work.[6] Why lean-mass gains don’t convert to force output — possibly fluid rather than contractile tissue, or lack of neural adaptation — is unresolved.
- Systemic vs local IGF-1. Secretagogues raise systemic IGF-1, but the repair-relevant signal may be locally, load-induced IGF-1/MGF. Whether systemic elevation adds meaningfully on top of a good training stimulus is unknown.
- Recovery has never been the primary endpoint. Studies measured GH, IGF-1, body composition, or strength — not time-to-recover, tendon-repair rate, or protein-synthesis kinetics after damage. The specific claim most people make about these blends is the least directly tested.
- Long-term safety. Chronic effects on glucose metabolism and the theoretical IGF-1/proliferation axis over years are not characterized for pulsatile secretagogue use.
- Dose, ratio, and timing optimization. The “best” ratio of GHRH-analog to ghrelin-mimetic, and the optimal timing relative to training and sleep, have not been established in controlled human work.
- Desensitization kinetics. Receptor biology predicts responsiveness could wane with over-frequent or tonic dosing, but the tachyphylaxis timeline for the specific blend in humans has not been measured.
Until controlled trials measure recovery endpoints for the actual blend, the scientifically defensible statement is narrow: Grow-H’s components can raise GH and IGF-1 in humans, those hormones activate pathways involved in muscle protein synthesis and repair, and therefore a recovery benefit is biologically plausible but unproven for the blend — and even the proven parts of the axis have a discouraging track record on strength and performance specifically.
Frequently Asked Questions
Is the Grow-H peptide blend FDA-approved for muscle recovery?
No. Grow-H is a research-only growth-hormone-secretagogue blend and is not an FDA-approved drug for muscle recovery, performance, or any other use. Its individual components are likewise not approved for these purposes in healthy people (tesamorelin, a related GHRH analog, is approved only for HIV-associated lipodystrophy). All recovery and performance discussion is educational and based on component-level and GH/IGF-1 physiology, not on trials of the finished product.
Does Grow-H actually build muscle or just raise IGF-1?
The evidence supports that its components raise GH and IGF-1 in humans, and sustained secretagogue exposure can modestly increase fat-free mass (about 1.1 kg in a two-year MK-677 trial). However, raising IGF-1 does not reliably translate into strength: the same body of GH-axis research shows lean-mass changes without measurable strength or performance gains. So “raises IGF-1” is well supported; “builds functional, performance-relevant muscle” is not established for the blend.
What is the difference between CJC-1295 with DAC and no DAC in a Grow-H blend?
CJC-1295 with DAC binds albumin and has a multi-day half-life (~5.8–8.1 days), producing continuous, tonic GH/IGF-1 elevation. CJC-1295 no DAC (modified GRF 1–29) has a ~30-minute half-life and produces a brief, timable pulse. Muscle-recovery-oriented blends usually use the no-DAC form paired with Ipamorelin so the combined signal stays pulsatile, which is thought to better mimic natural GH physiology than sustained elevation.
Why combine CJC-1295 with Ipamorelin instead of using one alone?
They act on different receptors. The GHRH analog (CJC-1295) raises GH pulse amplitude and supports GH synthesis via cAMP/PKA; Ipamorelin acts on the ghrelin receptor (GHS-R1a) to trigger release via calcium and to suppress somatostatin, the main inhibitory brake. Together they produce a larger, cleaner GH pulse than either alone, and Ipamorelin’s selectivity avoids the cortisol and prolactin spikes seen with older GHRPs.
Is there any human trial of the Grow-H blend specifically?
No published clinical trial has tested the assembled Grow-H (CJC-1295 + Ipamorelin) blend for muscle recovery, hypertrophy, or performance. The human data that exist are on the components individually and stop at GH/IGF-1 secretion and body-composition endpoints. Any performance claim for the branded blend is an inference across several steps, not a directly measured outcome.
Does Grow-H affect blood sugar or insulin sensitivity?
Potentially yes. GH is a counter-regulatory hormone that induces insulin resistance as part of its normal role, and sustained GH-secretagogue exposure has been shown to raise fasting glucose and worsen insulin sensitivity in long-term human data (the MK-677 trial). This is a key metabolic caveat and one reason brief pulsatile signaling is generally viewed more cautiously-favorably than continuous, tonic elevation of the GH/IGF-1 axis.
Can Grow-H improve tendon or connective-tissue recovery?
It is mechanistically plausible but unproven for the blend. GH and IGF-1 stimulate collagen synthesis in tendon and other extracellular-matrix tissue, which is why GH-axis manipulation is studied in connective-tissue repair contexts. However, no controlled study has measured tendon or connective-tissue recovery outcomes for the Grow-H blend, so this remains a theoretical benefit derived from GH/IGF-1 biology rather than a demonstrated result.
Why does GH raise lean mass but not strength?
This paradox appears consistently in the research. Part of the lean-mass increase attributed to GH reflects fluid retention and non-contractile tissue rather than added functional muscle, and strength depends heavily on neural adaptation and training stimulus that GH does not supply. The most rigorous systematic review of GH and athletic performance found increased lean body mass with no improvement in strength or exercise capacity, and some evidence of impaired capacity and more adverse events.
Why is Ipamorelin preferred over stronger GHRPs like hexarelin or GHRP-6?
Potency is not the only variable that matters. GHRP-6 strongly stimulates appetite and raises cortisol and prolactin; hexarelin is the most potent GHS-R1a agonist of the classic peptides but is comparatively prone to receptor desensitization with continued use and has additional CD36-mediated cardiac binding. Ipamorelin was engineered to retain GHRH-comparable GH-releasing potency while shedding the off-target cortisol/prolactin activation, giving a cleaner endocrine profile at GH-releasing doses. In a blend intended for repeated use, that selectivity and lower desensitization tendency generally outweigh raw potency.
Does the body build tolerance to Grow-H over time?
Receptor pharmacology predicts that continuous or excessive stimulation of the GHRH and ghrelin receptors can cause desensitization, internalization, and a blunted GH response — a general property of G-protein-coupled receptors. Intermittent, pulsatile dosing with short-acting components is thought to preserve responsiveness better than sustained exposure, which is one reason the no-DAC (short-acting) CJC-1295 form is favored over the multi-day DAC form. However, the specific desensitization kinetics of the assembled Grow-H blend in humans have not been characterized in controlled studies, so any tolerance timeline is inferred from receptor biology rather than measured for the product.
References
- Giustina A, Mazziotti G, Canalis E. Growth hormone, insulin-like growth factors, and the skeleton. Endocrine Reviews. 2008;29(5):535–559. https://academic.oup.com/edrv/article-abstract/29/5/535/2354995
- Teichman SL, Neale A, Lawrence B, Gagnon C, Castaigne JP, Frohman LA. Prolonged stimulation of growth hormone (GH) and insulin-like growth factor I secretion by CJC-1295, a long-acting analog of GH-releasing hormone, in healthy adults. J Clin Endocrinol Metab. 2006;91(3):799–805. https://pubmed.ncbi.nlm.nih.gov/16352683/
- Møller N, Jørgensen JOL. Effects of growth hormone on glucose, lipid, and protein metabolism in human subjects. Endocrine Reviews. 2009;30(2):152–177. https://pubmed.ncbi.nlm.nih.gov/19240267/
- Raun K, Hansen BS, Johansen NL, et al. Ipamorelin, the first selective growth hormone secretagogue. Eur J Endocrinol. 1998;139(5):552–561. https://pubmed.ncbi.nlm.nih.gov/9849822/
- Velloso CP. Regulation of muscle mass by growth hormone and IGF-I. Br J Pharmacol. 2008;154(3):557–568. https://bpspubs.onlinelibrary.wiley.com/doi/10.1038/bjp.2008.153
- Liu H, Bravata DM, Olkin I, et al. Systematic review: the effects of growth hormone on athletic performance. Ann Intern Med. 2008;148(10):747–758. https://www.acpjournals.org/doi/10.7326/0003-4819-148-10-200805200-00215
- Nass R, Pezzoli SS, Oliveri MC, et al. Effects of an oral ghrelin mimetic on body composition and clinical outcomes in healthy older adults: a randomized trial. Ann Intern Med. 2008;149(9):601–611. https://www.acpjournals.org/doi/10.7326/0003-4819-149-9-200811040-00003
- Alba M, Fintini D, Sagazio A, et al. Once-daily administration of CJC-1295, a long-acting growth hormone-releasing hormone (GHRH) analog, normalizes growth in the GHRH knockout mouse. Am J Physiol Endocrinol Metab. 2006;291(6):E1290–E1294. https://journals.physiology.org/doi/full/10.1152/ajpendo.00201.2006
- Holst B, Cygankiewicz A, Rosenkilde MM, Ankersen M, Schwartz TW. High constitutive signaling of the ghrelin receptor—identification of a potent inverse agonist. Mol Endocrinol. 2003;17(11):2201–2210. https://dx.doi.org/10.1210/me.2003-0069
- Camanni F, Ghigo E, Arvat E. Growth hormone-releasing peptides and their analogs. Front Neuroendocrinol. 1998;19(1):47–72. https://pubmed.ncbi.nlm.nih.gov/9465289/