The central research question behind the Glow peptide blend molecular mechanism is deceptively simple to state and genuinely difficult to answer: can a defined combination of signal and copper-binding peptides shift the behaviour of dermal fibroblasts toward a younger, more synthetically active state, and if so, through which intracellular pathways? In laboratory settings, bioactive peptides can bind cell-surface receptors, trigger intracellular signalling cascades, and modulate the transcription of genes that govern collagen, elastin, and glycosaminoglycan production. This article examines those proposed mechanisms in depth while remaining explicit about a crucial caveat: almost all of the supporting evidence is component-level and preclinical, drawn from fibroblast cultures, three-dimensional skin equivalents, and ex vivo tissue rather than from human clinical trials of the blend itself.
Research published in the American Journal of Pathology[1] demonstrates that fibroblasts from chronologically aged skin show reduced collagen production, driven both by intrinsic changes in fibroblast function and by defective mechanical stimulation from a fragmented matrix. This dual deficit — cells that make less collagen, embedded in a scaffold that no longer stretches them — frames the biological problem that peptide research aims to interrogate. Understanding how peptide signalling might, in a controlled model, restore fibroblast responsiveness and enhance matrix protein transcription is the mechanistic heart of the question.
Dosage Peptide highlights the importance of peptide purity, analytical characterization, documentation, and reproducibility in aesthetic research. These principles support controlled investigations of fibroblast signaling, extracellular matrix biology, and dermal regeneration while promoting methodological consistency and reliable interpretation of experimental findings in laboratory research settings.
What Is the Glow Peptide Blend, and What Does Its Evidence Actually Cover?
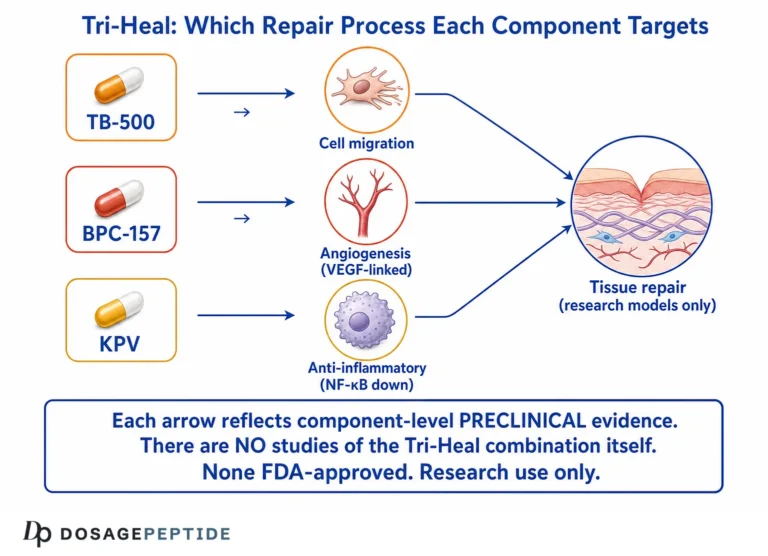
The Glow peptide blend, as offered for laboratory research, is a multi-component formulation that pairs a copper-binding signal peptide with regenerative peptides studied in tissue-repair contexts. As typically composed, the blend centres on GHK-Cu (glycyl-L-histidyl-L-lysine complexed with copper), the peptide most directly associated with dermal collagen and glycosaminoglycan signalling, and is often combined with BPC-157 and TB-500 (a thymosin beta-4 fragment), two peptides characterised almost entirely in wound-healing and angiogenesis models. The rationale for combining them is mechanistic complementarity: GHK-Cu is studied for direct matrix gene modulation, while BPC-157 and TB-500 are studied for angiogenic and cell-migration effects that, in principle, could support the tissue environment in which fibroblasts operate.
It is essential to be precise about evidentiary status before describing any mechanism. There are no human clinical trials of the Glow blend as a combined product. What exists is component-level evidence: GHK-Cu has decades of in-vitro and some in-vivo animal data plus cosmetic-grade topical use; BPC-157 and TB-500 have substantial preclinical (mostly rodent) literature but minimal human data and no regulatory approval for aesthetic indications. GHK-Cu itself is used as a cosmetic ingredient, not as an approved drug. Consequently, everything that follows about the Glow peptide blend molecular mechanism should be read as a synthesis of what each component does in laboratory systems, not as proof that the blend produces those outcomes in humans. This distinction is not a disclaimer — it is the accurate scientific picture.
Researchers examining reconstitution parameters and comparative single-agent behaviour often consult the Glow 70 mg vial research reference alongside standalone copper-peptide references such as the GHK-Cu 50 mg vial reference, since isolating the copper-peptide component is the only way to attribute observed effects to a specific molecule rather than to the blend as a whole.
Component overview and evidence tier
| Component | Class / origin | Primary studied mechanism | Strongest evidence tier |
|---|---|---|---|
| GHK-Cu | Copper-binding tripeptide (endogenous, declines with age) | Gene modulation, collagen/GAG synthesis, MMP–TIMP balance | In-vitro + some in-vivo animal; cosmetic topical use |
| BPC-157 | Synthetic gastric pentadecapeptide | Angiogenesis (VEGFR2), cytoprotection, cell migration | Preclinical rodent only; no aesthetic human trials |
| TB-500 | Fragment of thymosin beta-4 (actin-binding protein) | G-actin sequestration, cell migration, angiogenesis | Preclinical; most mechanistic data on parent Tβ4 |
| Glow blend (combined) | Multi-peptide research formulation | Proposed additive matrix + vascular support | No direct trials; inference from components only |
Why Does Aging Dermal Biology Set the Stage for Peptide Research?
To interpret any proposed peptide mechanism, one must first understand what changes in aging dermis, because peptides are hypothesised to counteract specific, well-characterised deficits. Skin aging is not a single process but a convergence of intrinsic (chronological) and extrinsic (largely photo-induced) damage that degrades the dermal extracellular matrix (ECM) and reprograms the cells that maintain it.
Chronoaging versus photoaging
Distinguishing intrinsic from extrinsic aging matters because they damage the matrix through partly different molecular routes, and a peptide mechanism may be more relevant to one than the other. Intrinsic (chronological) aging is driven by time-dependent processes — accumulating cellular senescence, mitochondrial decline, shortened telomeres, and a slow drop in growth-factor responsiveness — producing an atrophic dermis with reduced but relatively orderly collagen. Extrinsic aging, dominated by chronic ultraviolet exposure, drives a distinct pathology: UV generates reactive oxygen species that activate AP-1 and NF-κB, sharply inducing MMP-1, MMP-3, and MMP-9 while suppressing new procollagen synthesis. The result is solar elastosis — a build-up of abnormal, disorganised elastotic material alongside fragmented collagen. Because peptide research targets both the synthesis deficit and the degradative excess, the copper-peptide rationale is often framed as most relevant where the MMP–ROS axis is dominant.
The dermal-epidermal junction and basement membrane
Structural decline is not confined to the mid-dermis. The dermal-epidermal junction (DEJ) — the undulating basement-membrane interface rich in type IV and type VII collagen, laminins, and anchoring fibrils — flattens with age, reducing the surface area for nutrient and signal exchange between epidermis and dermis and weakening mechanical coupling. A flattened DEJ increases fragility and contributes to the appearance of thin, crepey skin. Any mechanism that supports basement-membrane component synthesis or fibroblast–keratinocyte crosstalk therefore has architectural relevance beyond fibrillar collagen alone, and 3D skin-equivalent models are valued precisely because they can reconstitute a rudimentary DEJ for study.
Fibroblast senescence and the loss of mechanical tension
Dermal fibroblasts are the workhorses of the dermis, synthesising collagen, elastin, fibronectin, and proteoglycans. With age, a growing fraction enter a senescent state: they stop dividing, adopt a senescence-associated secretory phenotype (SASP) that releases inflammatory cytokines and matrix-degrading enzymes, and downregulate collagen synthesis. The Varani work is pivotal here because it isolated two contributions: fibroblasts from older donors intrinsically produce less type I procollagen, and the aged matrix provides less mechanical stimulation to the cells that remain.[1] Fibroblasts sense the stiffness and stretch of their surroundings through integrins and mechanotransduction; when the collagen scaffold fragments, cells collapse, lose tension, and further reduce synthetic output — a self-reinforcing decline.
Collagen I/III decline and elastin fragmentation
Type I collagen provides the dominant tensile strength of the dermis, while type III collagen (more abundant in younger and wound-healing skin) contributes to the finer reticular network. The ratio of type III to type I shifts with age, and total collagen density falls. Simultaneously, the elastic fibre network — elastin cross-linked with fibrillin microfibrils — becomes fragmented and disorganised, and adult fibroblasts largely lose the robust elastogenesis seen in development. Because mature elastin turns over extremely slowly, damage accumulates and recoil is progressively lost, manifesting as laxity.
Glycosaminoglycan loss and reduced hydration
Glycosaminoglycans (GAGs) such as hyaluronic acid and the proteoglycan decorin fill the interfibrillar space, retain water, and organise collagen fibril spacing. Their decline reduces dermal hydration and turgor and alters how collagen fibrils assemble. GHK-Cu is of particular research interest here because it has been reported to stimulate not only collagen but also dermatan sulfate, chondroitin sulfate, and decorin, meaning its proposed action touches multiple ECM compartments at once rather than collagen alone.[6]
The MMP–TIMP imbalance
Matrix metalloproteinases (MMPs) degrade ECM as part of normal remodeling; their tissue inhibitors (TIMPs) restrain that activity. Ultraviolet exposure and inflammatory signalling drive MMP-1 (collagenase), MMP-3 (stromelysin), and MMP-9 upward, tilting the balance toward net degradation. A central hypothesis in dermal peptide research is that beneficial peptides do not merely add new collagen but rehabilitate the synthesis-to-degradation ratio — increasing structural protein output while damping excess MMP activity. Collagen-derived peptides, for example, have been shown in a physiologically crowded fibroblast model to increase elastin synthesis while significantly inhibiting MMP-1 and MMP-3 release.[7]
| Hallmark of dermal aging | Molecular change | Functional consequence | Peptide-research target? |
|---|---|---|---|
| Fibroblast senescence | SASP, reduced procollagen mRNA | Lower synthetic capacity | Yes – restore responsiveness (in-vitro) |
| Loss of mechanical tension | Integrin/mechanotransduction decline | Cell collapse, further synthesis drop | Indirect – via matrix rebuild |
| Collagen I/III decline | Reduced COL1A1/COL3A1 expression | Reduced tensile strength | Yes – transcriptional upregulation |
| Elastin fragmentation | Disorganised elastic fibres, low elastogenesis | Loss of recoil, laxity | Partial – elastin synthesis signals |
| GAG / proteoglycan loss | Less hyaluronan, decorin | Reduced hydration and fibril order | Yes – GHK-Cu multi-GAG signalling |
| MMP–TIMP imbalance | ↑ MMP-1/-3, relative ↓ TIMP | Net matrix degradation | Yes – rebalancing hypothesis |
What Molecular Pathways Regulate Dermal Regeneration?

Dermal regeneration is governed by tightly coordinated intracellular signalling pathways that regulate fibroblast proliferation, extracellular matrix deposition, and structural protein turnover. According to a 2024 review in Applied Sciences, cosmetic and bioactive peptides are grouped mechanistically into signal peptides, carrier peptides, enzyme-inhibitor peptides, and neurotransmitter-inhibitor peptides — and the copper-carrier and signal categories are those most relevant to collagen biology.[3] Below, each core pathway is examined for how it is proposed to translate a peptide signal into a matrix outcome. It is worth stressing that pathway assignments for the blend are inferred from single-peptide and general fibroblast studies; direct pathway mapping for the combined Glow blend has not been published.
TGF-β/Smad: the master collagen switch
Transforming growth factor-beta (TGF-β) is the dominant pro-fibrotic and pro-collagen signal in the dermis. When TGF-β binds its type II receptor, it recruits the type I receptor, which phosphorylates receptor-regulated Smads (Smad2/3); these partner with Smad4 and translocate to the nucleus to activate transcription of COL1A1, COL1A2, COL3A1, fibronectin, and connective-tissue growth factor. The inhibitory Smad7 provides negative feedback. In aged and photodamaged skin, TGF-β/Smad signalling is blunted, contributing to reduced collagen output. Peptides that can enhance or sensitise this axis — directly or by improving fibroblast health — are therefore of high mechanistic interest, because TGF-β/Smad sits closest to the collagen genes themselves.
PI3K/Akt: survival, proliferation, and protein synthesis
The phosphoinositide 3-kinase/Akt pathway integrates growth-factor and integrin signals to promote fibroblast survival, proliferation, and anabolic protein synthesis via downstream effectors including mTOR. In dermal models, activation of PI3K/Akt supports fibroblast viability and migration into a wound or remodeling zone and sustains the translational machinery needed for high-output collagen production. Copper-peptide and collagen-peptide studies frequently report increased fibroblast proliferation, a readout consistent with PI3K/Akt engagement.[7] BPC-157 has also been linked in preclinical work to Akt/PI3K survival signalling as part of its cytoprotective profile.
MAPK/ERK: growth control and remodeling dynamics
The mitogen-activated protein kinase (MAPK) cascade — particularly the ERK1/2 branch — transduces mitogenic signals into proliferation and modulates the expression of remodeling enzymes. ERK signalling is a double-edged sword in skin: it drives proliferation and can support repair, but sustained ERK activation downstream of UV-induced stress also upregulates AP-1 (c-Jun/c-Fos), which transcriptionally boosts MMPs and suppresses collagen. A favourable peptide mechanism would ideally bias MAPK output toward proliferation and organised remodeling rather than toward the AP-1–driven degradative program, illustrating why context and dose dominate outcomes in these systems.
NF-κB: the inflammatory brake on collagen
Nuclear factor kappa-B (NF-κB) is the central hub for inflammatory transcription. Chronic NF-κB activation — a feature of “inflammaging” and the SASP — sustains pro-inflammatory cytokines (IL-1, IL-6, TNF-α) that induce MMPs and suppress matrix synthesis. One of the most cited properties attributed to GHK-Cu in the gene-expression literature is suppression of NF-κB–associated signalling, positioning it as an anti-inflammatory modulator that could indirectly protect collagen by lowering the degradative tone of the tissue.[2]
Nrf2/antioxidant response: defending newly made matrix
Nuclear factor erythroid 2–related factor 2 (Nrf2) is the master regulator of the antioxidant response element (ARE), driving expression of superoxide dismutase, catalase, glutathione enzymes, and heme oxygenase-1. Reactive oxygen species (ROS) both damage collagen directly and activate the MMP-inducing AP-1/NF-κB programs, so antioxidant capacity is protective of matrix integrity. Copper is a cofactor for Cu/Zn-superoxide dismutase, and GHK-Cu is studied for its capacity to influence antioxidant defences — a plausible route by which the copper-peptide component could stabilise freshly synthesised collagen against oxidative breakdown.[6]
| Signaling pathway | Key effectors | Downstream ECM effect | Proposed peptide role |
|---|---|---|---|
| TGF-β/Smad | Smad2/3–Smad4; CTGF | ↑ COL1A1, COL1A2, COL3A1, fibronectin | Enhance/sensitise collagen transcription |
| PI3K/Akt/mTOR | Akt, mTOR | Fibroblast survival, proliferation, translation | Sustain synthetic cell population |
| MAPK/ERK | ERK1/2, AP-1 | Proliferation vs. MMP induction (context-dependent) | Bias toward organised remodeling |
| NF-κB | p65/p50, IL-1/-6, TNF-α | ↑ MMPs, ↓ matrix synthesis when chronic | Damp inflammatory degradation |
| Nrf2/ARE | SOD, catalase, HO-1, glutathione | Protect collagen from ROS/MMP induction | Stabilise newly formed matrix |
When these pathways decline with age or oxidative stress, dermal structure weakens. The mechanistic thesis of peptide research is that specific sequences can recalibrate one or more of these signalling systems under controlled conditions — but demonstrating that a blend does so coherently, and reproducibly, is a much higher bar than showing that a single peptide nudges one pathway in a dish.
Pathway crosstalk and the AP-1 node
These pathways do not operate in isolation; they converge and antagonise one another, which is why single-pathway thinking often misleads. The activator protein-1 (AP-1) transcription factor, assembled from c-Jun and c-Fos, is a critical convergence point: it is activated downstream of MAPK/ERK and stress signalling, it induces MMP transcription, and it directly antagonises TGF-β/Smad signalling by blocking Smad-driven collagen transcription. This means a peptide that raises collagen mRNA and one that lowers MMP output may in fact be acting on two ends of the same regulatory tug-of-war. Similarly, NF-κB and Nrf2 are reciprocally regulated — boosting antioxidant tone via Nrf2 can indirectly dampen NF-κB–driven inflammation. A credible account of the Glow peptide blend molecular mechanism therefore has to consider net signalling balance rather than isolated arrows on a diagram, which is one reason blend behaviour is hard to predict from single-component data.
How Are Peptides Sensed at the Cell Surface?
Before any intracellular cascade fires, the peptide must be detected by the cell — and the mechanism of detection differs by peptide class. Understanding this step clarifies why some peptides are called “signal” peptides, others “carrier” peptides, and why the distinction shapes what evidence is even possible to gather. The 2024 Applied Sciences review organises cosmetic peptides into exactly these functional categories, each with a different point of molecular action.[3]
Matrikines and the fragment-signalling concept
Many dermally active peptides are matrikines — short fragments released by partial enzymatic breakdown of ECM proteins such as collagen, elastin, and fibronectin. These fragments act as feedback signals: their appearance tells fibroblasts that matrix turnover is occurring and, in effect, prompts a synthetic or remodeling response. This is the conceptual basis for why collagen-derived peptides stimulate the very fibroblasts that make collagen — they mimic the endogenous fragments generated during natural remodeling.[4] The Linus Pauling Institute similarly notes that naturally occurring bioactive skin peptides are generated by enzymatic processing of endogenous proteins, framing exogenous peptides as an attempt to supply that same signal.[5]
Integrins, mechanotransduction, and receptor engagement
Fibroblasts read their environment largely through integrins — transmembrane receptors that bind ECM ligands (including RGD motifs in fibronectin) and transduce both chemical and mechanical information inward via focal adhesion kinase (FAK) into PI3K/Akt and MAPK/ERK. This is why the loss of mechanical tension in aged skin is so consequential: it is a loss of integrin signalling. Some peptides are proposed to engage or sensitise these adhesion pathways, while carrier peptides such as GHK-Cu work substantially through metal-ion delivery rather than classical receptor binding. The precise receptor(s) for GHK-Cu remain incompletely defined, and this mechanistic uncertainty is itself an honest limitation: much is known about GHK-Cu’s downstream effects and comparatively less about the exact proximal event that initiates them.
Why detection mechanism constrains the evidence
Because carrier peptides act partly through ion delivery and signal peptides through fragment-mimicry, the assays that best capture each differ, and effects can be exquisitely concentration-dependent — GHK-Cu’s collagen stimulation, for instance, is reported in a narrow low-nanomolar window rather than rising monotonically with dose.[6] A blend combines molecules with different optimal windows and different detection mechanisms, which further complicates predicting a single coherent dose-response — another reason the combined blend is harder to characterise than any one component.
How Does GHK-Cu Work at the Molecular Level?
GHK-Cu is the component around which the Glow blend’s dermal rationale is built, and it is also the component with the deepest mechanistic literature. GHK (glycyl-L-histidyl-L-lysine) is a naturally occurring tripeptide present in plasma, saliva, and urine whose concentration declines markedly with age — a fact often used to frame it as a signal the aging body loses. Its defining biochemical feature is a high affinity for copper(II) ions; the histidine imidazole and terminal amine coordinate Cu²+ to form the GHK-Cu complex, and much of its biological activity is attributed to this copper-carrying capacity rather than to the bare peptide.
Copper delivery as a signalling event
Copper is a required cofactor for lysyl oxidase (which cross-links collagen and elastin), for Cu/Zn-superoxide dismutase (antioxidant defence), and for several enzymes involved in angiogenesis. By chelating and delivering copper in a bioavailable form, GHK-Cu is proposed to support these copper-dependent processes locally. This is a mechanistically attractive story because it links a single molecule to structural cross-linking, antioxidant capacity, and vascular support simultaneously. In fibroblast culture, GHK-Cu has long been reported to stimulate collagen synthesis at very low (picomolar to nanomolar) concentrations, an observation consistent with a signalling rather than nutritive role.
Broad gene modulation — and how to read it honestly
The most striking claim in the GHK literature is that the peptide can modulate the expression of thousands of human genes. This derives from analyses using the Broad Institute’s Connectivity Map, a database of transcriptional responses in cultured cell lines exposed to compounds. Pickart and Margolina report that GHK influences a very large number of genes — broadly upregulating those tied to tissue repair, DNA repair, and antioxidant defence while downregulating a set tied to inflammation and cancer-associated processes.[2] The honest framing is critical here: these are in-vitro transcriptomic signatures in cell lines, not demonstrations that topical or injected GHK-Cu reprograms human dermis in vivo. The gene data describe a plausible mechanistic potential; they are not clinical proof of an anti-aging outcome, and the authors themselves present them as a mechanistic hypothesis grounded in cell-line data.[6]
Dual action on synthesis and degradation
A recurring theme is that GHK-Cu appears to act on both sides of the remodeling equation: stimulating synthesis of collagen and multiple glycosaminoglycans (including decorin, dermatan sulfate, and chondroitin sulfate) while also modulating the activity of MMPs and their TIMP inhibitors.[6] This “remodel, don’t just add” behaviour is mechanistically distinct from simply flooding a culture with a growth factor, and it is part of why GHK-Cu is treated as a signal peptide rather than a bulk substrate. Researchers isolating this behaviour frequently compare vial strengths using the GHK-Cu 100 mg vial reference to hold copper-peptide concentration constant across experimental arms.
| GHK-Cu reported action | Molecular basis | ECM/skin readout | Evidence context |
|---|---|---|---|
| Collagen stimulation | Signal to fibroblasts; TGF-β-associated | ↑ type I / type III collagen | In-vitro, low nM |
| GAG / proteoglycan synthesis | Decorin, dermatan/chondroitin sulfate | Improved hydration, fibril order | In-vitro / ex vivo |
| MMP–TIMP modulation | Balances proteinase vs. inhibitor | Controlled remodeling | In-vitro |
| Antioxidant support | Copper for SOD; Nrf2-linked | Protection of matrix from ROS | In-vitro / mechanistic |
| Anti-inflammatory | NF-κB-associated suppression | Lower degradative tone | Cell-line transcriptomics |
| Broad gene modulation | Connectivity Map signature | Repair-biased expression | Cell-line, hypothesis-generating |
Copper homeostasis and research safety considerations
The same copper chemistry that makes GHK-Cu interesting also imposes constraints that responsible research must respect. Copper is essential but redox-active: in excess or in the wrong context it can catalyse Fenton-type reactions that generate hydroxyl radicals, potentially damaging the very matrix the peptide is meant to support. Cells maintain tight copper homeostasis through chaperones and transporters (such as ATP7A/B and the CTR1 importer), and a benefit of the peptide-bound form is that copper is delivered in a coordinated, buffered state rather than as free ion. This is why concentration control is not a formality in copper-peptide research — the difference between a pro-synthetic signal and an oxidative insult can be a matter of dose and local copper load. In experimental design, this argues for narrow concentration ranges, characterised copper content, and antioxidant-status readouts alongside collagen endpoints. It also underscores that “more is better” reasoning is inappropriate for a copper carrier; the reported biphasic, window-dependent activity of GHK-Cu is consistent with this biology.[6]
What Do BPC-157 and TB-500 Contribute at the Component Level?
If GHK-Cu supplies the matrix-transcription rationale, BPC-157 and TB-500 supply a tissue-environment rationale — and their evidence base is more preliminary still. Both are studied predominantly in wound-healing and musculoskeletal-repair models, and neither is an approved drug; indeed, BPC-157 is explicitly flagged by anti-doping and regulatory bodies as an unapproved substance. Their inclusion in a dermal “glow” blend is a reasonable mechanistic hypothesis, not a validated dermatological indication.
BPC-157: angiogenesis and cytoprotection
BPC-157 is a synthetic pentadecapeptide derived from a protein found in gastric juice. In rodent models it accelerates healing across tendon, ligament, muscle, and gut tissue, with the mechanistic story centred on angiogenesis (upregulation of VEGFR2 signalling and increased vascular density in repair zones), nitric-oxide pathway modulation, and FAK-paxillin–dependent endothelial migration.[8] The dermal relevance is indirect: better local perfusion could, in principle, support fibroblast metabolism and matrix production. But the review literature is unambiguous that this evidence is preclinical — the pathway’s equivalence in human skin has not been established, and human trials for these effects have not been conducted.[8]
TB-500: actin dynamics and cell migration
TB-500 is a synthetic fragment corresponding to the active region of thymosin beta-4 (Tβ4), one of the most abundant actin-binding proteins in mammalian cells. Its defining mechanism is G-actin sequestration: by binding monomeric actin and regulating the G-actin/F-actin equilibrium, it influences cytoskeletal remodeling that underlies cell migration, angiogenesis, and tissue repair. A recent scoping review of Tβ4 and TB-500 catalogues repair mechanisms including endothelial and progenitor cell migration, tube formation, inflammatory modulation, and ECM remodeling — while noting that most robust mechanistic data pertain to the parent Tβ4 rather than to TB-500 specifically, and that the evidence base is preclinical.[9] For dermal research, the plausible contribution is enhanced fibroblast and endothelial migration into a remodeling zone, complementing GHK-Cu’s synthetic signal.
The combined logic is that GHK-Cu drives matrix gene expression, TB-500 supports the cell migration needed to populate and organise that matrix, and BPC-157 supports the vascular supply that sustains it. This is a coherent hypothesis on paper. It is not, at present, a demonstrated synergy: no controlled study has shown that the three peptides together outperform GHK-Cu alone in a dermal model, and additive or even antagonistic interactions cannot be assumed without direct testing.
What Research Reveals About the Glow Peptide Blend and Extracellular Matrix Remodeling
Scientific literature demonstrates that signal and copper-binding peptides can stimulate fibroblasts to increase synthesis of structural proteins and matrix components. The 2024 Applied Sciences review confirms that bioactive peptides can enhance collagen, elastin, fibronectin, and proteoglycan production in dermal models and organises the mechanisms by peptide class.[3] Translating those transcriptional signals into structural readouts is where mechanism meets measurement.
Collagen density: COL1A1 and COL3A1
Upregulation of COL1A1 and COL3A1 transcription increases procollagen output, which — after post-translational hydroxylation, secretion, cleavage, and lysyl oxidase–mediated cross-linking — raises fibrillar collagen density and the tensile strength of dermal equivalents. A recombinant collagen α-1 type I–derived protein was shown to induce fibroblast proliferation and collagen biosynthesis, as well as migration and elastin production, in human dermal fibroblasts, illustrating how collagen-family signals feed back to stimulate their own network.[4]
Elastin network and fibronectin
Elastogenesis in adult fibroblasts is limited, so any signal that increases elastin (ELN) expression and supports fibrillin scaffolding is mechanistically valuable for recoil. Collagen-derived peptides have been reported to significantly increase fibroblast elastin synthesis while simultaneously inhibiting elastin degradation, alongside reduced MMP-1 and MMP-3 release.[7] Fibronectin, an early provisional-matrix glycoprotein, guides fibroblast adhesion and organises subsequent collagen assembly; its upregulation is part of a coordinated pro-synthetic program rather than an isolated effect.
Proteoglycans and the MMP–TIMP set-point
Proteoglycans such as decorin regulate collagen fibril diameter and spacing; their synthesis is one of the distinguishing features attributed to GHK-Cu.[6] Just as important as making matrix is not over-degrading it: the recurring in-vitro finding across peptide studies is a shift in the MMP–TIMP set-point toward preservation, so that newly synthesised collagen and elastin persist rather than being cleared. This synthesis-plus-protection pattern is the strongest unifying mechanistic theme in the dermal peptide literature.
| Component | Target pathway / signal | ECM readout studied | Representative in-vitro evidence |
|---|---|---|---|
| GHK-Cu | TGF-β-linked, Nrf2, NF-κB, copper enzymes | Collagen, GAGs, decorin; MMP–TIMP balance | Fibroblast synthesis at low nM; gene signatures[6] |
| Collagen-derived peptides (model) | Proliferation, elastin signalling | ↑ elastin, ↓ MMP-1/-3 | Crowded HDF culture[7] |
| hCOL1A1-derived protein | Fibroblast proliferation/migration | Collagen + elastin production | Human dermal fibroblasts[4] |
| TB-500 (Tβ4) | Actin dynamics, migration, angiogenesis | Cell migration, ECM remodeling | Preclinical scoping review[9] |
| BPC-157 | VEGFR2 angiogenesis, cytoprotection | Vascularity, granulation, collagen content | Rodent repair models[8] |
How Does the Glow Peptide Blend Influence Collagen and Elastin Gene Expression?
At the transcriptional level, the proposed influence begins when a peptide signal is sensed by fibroblasts — through cell-surface interactions and, for copper peptides, through copper-dependent enzyme support — and propagates via the TGF-β/Smad and PI3K/Akt cascades to raise mRNA for collagen α-chains and elastin-associated proteins. Quantitative studies in human dermal fibroblasts show that collagen-family and collagen-derived peptides can increase collagen biosynthesis, proliferation, migration, and elastin production, confirming that peptide signalling is capable of modulating dermal gene expression in controlled systems.[4]
From transcription to a stable fibre
Raising COL1A1/COL3A1 mRNA is only the first step. Procollagen must be hydroxylated (a vitamin-C– and iron-dependent step), assembled into triple helices, secreted, cleaved to tropocollagen, and cross-linked by lysyl oxidase — a copper enzyme. This is where the copper-delivery mechanism of GHK-Cu becomes conceptually elegant: the same molecule that is proposed to signal for more collagen also delivers the copper cofactor needed to cross-link and stabilise it. Whether this coupling operates efficiently in intact human dermis, as opposed to a culture dish, remains an open experimental question rather than a settled fact.
Protecting the product
Copper-binding peptides are also studied for their regulation of oxidative-stress pathways and upregulation of antioxidant enzymes, which would protect newly synthesised collagen fibres from ROS-mediated degradation.[2] This dual mechanism — stimulating synthesis while reducing breakdown — is the mechanistic case for why a copper-peptide-centred blend might, in principle, strengthen dermal architecture more durably than a synthesis-only stimulus. Researchers formulating comparators frequently consult the peptide reconstitution guide to keep solvent, concentration, and storage variables constant, since these directly affect peptide stability and therefore any measured gene-expression response.
How Does Research Support the Glow Peptide Blend in Topical and Injectable Studies?
How a peptide reaches the dermis fundamentally shapes what mechanism can be observed. The Linus Pauling Institute notes that bioactive peptides can be generated by enzymatic processing of endogenous skin proteins and that topical application of such peptides is used primarily for cosmetic purposes, while cautioning that clinical trials are needed to establish efficacy and safety — an important, honest anchor for any delivery discussion.[5]
Topical delivery and the stratum corneum barrier
The stratum corneum is a formidable barrier to hydrophilic peptides. Research formulations evaluate low peptide concentrations for epidermal penetration and dermal gene activation, using encapsulation (liposomes, nanoparticles) and permeation enhancers to improve delivery. A key research question is whether an intact peptide reaches viable fibroblasts in sufficient quantity to trigger the signalling described above, or whether observed surface effects reflect other mechanisms. Penetration is size- and charge-dependent, and small peptides like GHK have an advantage over larger ones.
Microneedle-assisted delivery
Microneedling creates transient microchannels that bypass the stratum corneum, allowing deeper peptide diffusion into the dermis. Beyond delivery, the controlled micro-injury itself triggers a wound-healing cascade (including TGF-β and growth-factor release), which can confound interpretation: an observed effect may reflect the needling stimulus, the peptide, or their interaction. Well-designed studies therefore include needling-only controls to isolate the peptide-specific mechanism.
Injectable research formulations
Injectable research formulations deliver peptides directly into dermal layers, enabling precise molecular interactions with fibroblast populations and controlled dose-response study. This route removes the penetration variable but introduces its own considerations — local concentration, clearance, and tissue distribution. Because dosing is central to reproducibility, researchers frequently model target concentrations with a reconstitution and dosage calculator before preparing experimental arms.
3D skin equivalents and ex vivo models
Peptide stability as a delivery variable
Delivery is not only about crossing a barrier; it is about whether an intact, active molecule survives to reach its target. Small peptides are vulnerable to enzymatic degradation by skin and serum peptidases, to oxidation, and to aggregation, and their activity can be lost during reconstitution, freeze-thaw cycling, or storage in unsuitable solvents. For copper peptides, the coordination state of the copper must also be preserved. These factors mean that two laboratories using nominally the same peptide can obtain different results simply because of handling, which is why documented reconstitution, controlled aliquoting, and cold-chain storage are treated as part of the experimental method rather than as afterthoughts. Standardising these variables is a prerequisite for attributing any observed gene-expression change to the peptide itself rather than to formulation drift.
Between simple monolayer culture and human trials sit two intermediate systems. Three-dimensional skin equivalents — fibroblast-populated dermal matrices topped with stratified keratinocytes — reconstruct much of the tissue architecture and allow simultaneous study of penetration and dermal response. Ex vivo human skin explants preserve the native matrix and cell populations for short-term study. These models are far more physiologically relevant than monolayers, yet they remain models: they lack circulation, immune trafficking, and the chronic timescales over which human dermal remodeling actually occurs. The gap between a promising 3D-equivalent result and a demonstrated human outcome is exactly the gap that keeps the Glow blend in the research, not the proven, category.
What Analytical Methods Measure Peptide-Induced Dermal Remodeling?
A mechanistic claim is only as strong as the assays behind it, and dermal peptide research draws on a stack of complementary methods spanning transcript, protein, and tissue levels. Triangulating across these — rather than relying on any single readout — is what distinguishes a convincing mechanism from an artefact.
Transcript-level: qPCR and RNA-seq
Quantitative PCR (qPCR) measures the relative abundance of specific mRNAs — COL1A1, COL3A1, ELN, fibronectin, MMPs, TIMPs — and is the standard first readout of a transcriptional effect. RNA sequencing broadens this to genome-wide expression, the approach conceptually behind the Connectivity Map signatures attributed to GHK.[2] A limitation to keep in view: elevated mRNA does not guarantee more functional protein, which is why protein-level confirmation is essential.
Protein-level: ELISA and Western blot
Enzyme-linked immunosorbent assay (ELISA) quantifies secreted proteins such as procollagen type I C-peptide, MMP-1, and MMP-3 in culture media, while Western blotting resolves protein amount and, using phospho-specific antibodies, the activation state of signalling nodes (phospho-Smad2/3, phospho-Akt, phospho-ERK). Together they connect a gene-expression change to actual protein output and to the pathway responsible.
Tissue-level: IHC/IF and hydroxyproline
Immunohistochemistry (IHC) and immunofluorescence (IF) visualise the spatial organisation of collagen, elastin, and fibronectin within a 3D equivalent or explant — distinguishing well-organised fibres from disordered deposition. The hydroxyproline assay quantifies total collagen biochemically, since hydroxyproline is a collagen-specific amino acid; it is a gold-standard bulk measure of collagen content. Picrosirius red staining under polarised light adds information on fibre thickness and cross-linking.
| Assay method | Level probed | Typical readout | Key caveat |
|---|---|---|---|
| qPCR | mRNA | Relative COL1A1/COL3A1/ELN/MMP expression | mRNA ≠ functional protein |
| RNA-seq | Transcriptome | Genome-wide expression signature | Cell-line data ≠ in-vivo outcome |
| ELISA | Secreted protein | Procollagen, MMP-1/-3 levels | Media only; not spatial |
| Western blot | Protein / phospho-state | p-Smad2/3, p-Akt, p-ERK | Semi-quantitative |
| IHC / IF | Tissue architecture | Fibre localisation and organisation | Antibody specificity |
| Hydroxyproline | Bulk biochemistry | Total collagen content | No fibre-type resolution |
Controls that make a result interpretable
The assays above only yield meaningful conclusions when embedded in disciplined controls, and this is where much low-quality peptide research falls down. A credible study includes a vehicle control (the solvent and, for copper peptides, an appropriate copper-only or peptide-only comparator to separate the metal’s contribution from the peptide’s), a dose-response series rather than a single concentration, and — for blends — single-component arms so that any additive effect can be detected against each part alone. Positive controls (for example, TGF-β or ascorbic acid for collagen synthesis) confirm the assay can detect a real change, while housekeeping-gene normalisation and biological replicates guard against technical artefacts. Blinded quantification of imaging endpoints reduces observer bias. When a copper-peptide result is reported without a copper-only control, or a blend result without single-agent arms, the data cannot distinguish the proposed mechanism from simpler explanations — a limitation that should temper how confidently any such finding is cited.
How Strong Is the Evidence for the Glow Peptide Blend, Really?
An honest evidence review requires separating three tiers of confidence. First, the biology of dermal aging (collagen decline, MMP imbalance, fibroblast senescence) is well established in humans and supported by primary pathology data.[1] Second, the mechanistic potential of the individual peptides is supported by in-vitro and preclinical studies of real quality — GHK-Cu’s collagen/GAG signalling and gene modulation,[2][6] collagen-peptide effects on elastin and MMPs,[7] and the angiogenic/migratory profiles of BPC-157 and TB-500.[8][9] Third — and this is where honesty matters most — the Glow blend as a combined, in-vivo, human intervention is unproven. There is no clinical trial of the blend, and cross-component synergy is hypothesised, not measured.
Key limitations to weigh
- Model-to-human gap: monolayers, 3D equivalents, and rodents do not reproduce human dermal timescales, circulation, or immunity.
- mRNA-to-outcome gap: transcriptomic signatures are hypothesis-generating, not proof of clinical benefit.
- Blend-specific data absent: effects are inferred from single peptides; interactions could be additive, redundant, or antagonistic.
- Delivery uncertainty: whether intact peptide reaches fibroblasts at active concentrations is formulation-dependent.
- Regulatory status: GHK-Cu is a cosmetic ingredient, not an approved drug; BPC-157 is an unapproved substance flagged by anti-doping bodies. None is approved for the effects discussed.
- Publication and quality variance: some component literature appears in lower-tier venues; primary, peer-reviewed sources should anchor any claim.
Reading component evidence without over-extrapolating
A common error in this space is transitive reasoning: because GHK-Cu stimulates collagen in fibroblasts, and because BPC-157 promotes angiogenesis in rodents, the blend must rejuvenate human skin. Each premise can be individually well-supported while the conclusion remains unproven, because the inference skips several real steps — delivery to the target cells at active concentration, preservation of each peptide’s optimal window within a shared formulation, absence of interference between components, and translation from a model system to intact, living human dermis over clinically meaningful timescales. The strongest primary sources here are careful about exactly this boundary: the GHK gene and pathway reviews present mechanistic potential grounded in cell and animal data,[2][6] and the BPC-157 and thymosin reviews explicitly emphasise the preclinical nature of the evidence and the need for human trials.[8][9] When a claim about the blend exceeds what these primary sources support, the extrapolation — not the underlying science — is where it fails. Prioritising peer-reviewed primary literature over marketing summaries, and matching each specific claim to a source that actually tested it, is the discipline that keeps a mechanistic discussion honest.
Read correctly, the Glow peptide blend molecular mechanism is a well-reasoned, component-supported hypothesis about how copper-peptide signalling plus angiogenic and migratory support could shift dermal fibroblast behaviour — a legitimate and interesting research question, but not a demonstrated clinical result.
How Does the Blend Compare With Individual Peptides?
A recurring practical question is whether a blend offers any mechanistic advantage over a single, better-characterised peptide such as GHK-Cu. The theoretical case for a blend is coverage of complementary mechanisms — synthesis (GHK-Cu), migration (TB-500), and vascular support (BPC-157). The theoretical case for a single agent is interpretability: with one variable, an observed effect can be attributed unambiguously, dose-response is clean, and results are reproducible. In rigorous research design, the single-peptide arm is almost always the more informative starting point, with blends studied only after each component’s behaviour is understood.
For this reason, comparative studies typically include isolated GHK-Cu arms at matched copper-peptide concentrations. Researchers frequently reference standalone copper-peptide parameters against the combined Glow blend reference to hold the copper-peptide fraction constant between a single-agent and a blend arm, isolating whether the additional peptides contribute measurable effect. Terminology across these designs can be consulted in the peptide research glossary. Until such head-to-head data exist, any claim that the blend outperforms GHK-Cu alone is speculative.
Advancing Molecular Skin Research with Rigor
Researchers working with multi-peptide dermal formulations repeatedly encounter the same obstacles: variability in peptide purity, inconsistent receptor activation across batches, delivery uncertainty, and insufficient mechanistic documentation. Each undermines reproducibility and complicates interpretation. Without validated purity data, characterised signalling readouts, and matched controls, drawing reliable conclusions about collagen or elastin enhancement is not possible — regardless of how compelling the underlying hypothesis appears. High-quality sourcing, transparent analytical characterization, and disciplined experimental design remain the prerequisites for turning a plausible mechanism into defensible data, and for keeping honest the line between what has been observed in a dish and what has been demonstrated in human skin.
Frequently Asked Questions
What is the Glow peptide blend?
The Glow peptide blend is a research-grade, multi-component formulation built around GHK-Cu, a copper-binding signal peptide studied for collagen and glycosaminoglycan signalling, and typically combined with BPC-157 and TB-500. It is used in laboratory settings to investigate fibroblast signalling and extracellular-matrix biology. It is not an approved drug, and no human clinical trials exist for the blend as a combined product.
What is the Glow peptide blend molecular mechanism, in brief?
Mechanistically, the components are proposed to act complementarily: GHK-Cu signals fibroblasts to upregulate collagen, GAG, and antioxidant pathways (via TGF-β/Smad, Nrf2, and NF-κB modulation) while delivering copper for cross-linking; TB-500 supports actin-driven cell migration; and BPC-157 supports angiogenesis. This is a component-level, largely in-vitro and preclinical hypothesis rather than a demonstrated human outcome.
Is the Glow blend clinically proven to rejuvenate skin?
No. There are no human clinical trials of the blend, and it is not approved for any aesthetic indication. The available evidence is component-level and comes from fibroblast cultures, 3D skin equivalents, ex vivo tissue, and animal models. These support a mechanistic hypothesis but do not establish efficacy or safety in humans, which would require controlled clinical study.
How does GHK-Cu differ from the other components?
GHK-Cu has the deepest dermal literature: it is studied for directly stimulating collagen and glycosaminoglycan synthesis, modulating the MMP–TIMP balance, and reprogramming gene expression in cell-line analyses. BPC-157 and TB-500, by contrast, are characterised mainly in wound-healing and angiogenesis models with minimal dermal-specific or human data, so their contribution to a “glow” effect is more speculative.
Does the blend influence oxidative-stress pathways?
The copper-binding component (GHK-Cu) is studied for modulating oxidative stress by supporting antioxidant enzymes such as superoxide dismutase and by influencing Nrf2-linked defences, since copper is a required SOD cofactor. In principle this could reduce reactive-oxygen-species–mediated collagen degradation and help stabilise newly formed matrix. These findings are from in-vitro and mechanistic studies, not human trials of the blend.
Which experimental models are used to study these mechanisms?
Common systems include monolayer human dermal fibroblast cultures, three-dimensional skin equivalents (fibroblast dermal matrices with stratified keratinocytes), and ex vivo human skin explants, plus rodent models for the angiogenic components. Each adds physiological relevance but none replicates the full human dermal environment, timescale, circulation, or immune context — a key reason results remain preclinical.
What analytical methods measure peptide-induced remodeling?
Researchers use qPCR and RNA-seq for collagen, elastin, and MMP gene expression; ELISA and Western blotting for secreted proteins and pathway phosphorylation; immunohistochemistry and immunofluorescence for matrix organisation; and the hydroxyproline assay for total collagen content. Triangulating across transcript, protein, and tissue levels — with matched controls — is what makes a mechanistic conclusion credible.
Why study a single peptide instead of the blend?
Single-peptide arms give clean, interpretable dose-response data and let an observed effect be attributed to one molecule. Blends introduce multiple simultaneous variables whose interactions may be additive, redundant, or antagonistic. Sound design characterises each component first — often with isolated GHK-Cu at matched concentration — before testing whether a blend adds measurable value.
References
- Varani, J., Dame, M. K., Rittié, L., Fligiel, S. E. G., Kang, S., Fisher, G. J., & Voorhees, J. J. (2006). Decreased collagen production in chronologically aged skin: roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. American Journal of Pathology, 168(6), 1861–1868.
- Pickart, L., & Margolina, A. (2018). Regenerative and protective actions of the GHK-Cu peptide in the light of the new gene data. International Journal of Molecular Sciences, 19(7), 1987.
- Waszkielewicz, A. M., & Mirosław, K. (2024). Peptides and their mechanisms of action in the skin. Applied Sciences, 14(24), 11495.
- Hwang, S. J., Kim, S. H., Seo, W.-Y., Jeong, Y., Shin, M. C., Ryu, D., Lee, S. B., Choi, Y. J., & Kim, K. (2021). Effects of human collagen α-1 type I-derived proteins on collagen synthesis and elastin production in human dermal fibroblasts. BMB Reports, 54(6).
- Linus Pauling Institute. Peptides and skin health. Micronutrient Information Center, Oregon State University.
- Pickart, L., Vasquez-Soltero, J. M., & Margolina, A. (2015). GHK peptide as a natural modulator of multiple cellular pathways in skin regeneration. BioMed Research International, 2015, 648108.
- Edgar, S., Hopley, B., Genovese, L., Sibilla, S., Laight, D., & Shute, J. (2018). Effects of collagen-derived bioactive peptides and natural antioxidant compounds on proliferation and matrix protein synthesis by cultured normal human dermal fibroblasts. Scientific Reports, 8, 10474.
- Gwyer, D., Wragg, N. M., & Wilson, S. L. (2019). Gastric pentadecapeptide body protection compound BPC 157 and its role in accelerating musculoskeletal soft tissue healing. Cell and Tissue Research, 377(2), 153–159.
- Thymosin beta-4 and TB-500 in tissue healing, regeneration, and musculoskeletal repair: a scoping review (2026). Applied Sciences, 16(12), 6202.